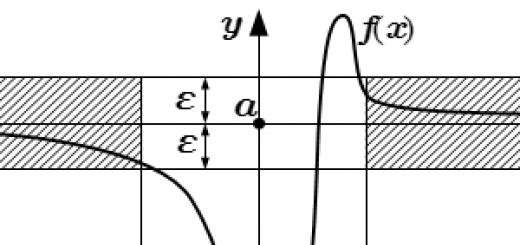
Спинний мозок характеризується вираженою сегментарною будовою, що відображає сегментарну будову тіла хребетних. Від кожного спинномозкового сегмента відходять дві пари вентральних та дорсальних корінців. Дорсальні коріння формують аферентні входи спинного мозку. Вони утворені центральними відростками волокон первинних аферентних нейронів, тіла яких винесені на периферію та знаходяться у спинномозкових гангліях. Вентральні коріння утворюють еферентні виходи спинного мозку. У них проходять аксони a та g-мотонейронів, а також прегангліонарних нейронів вегетативної. нервової системи. Такий розподіл аферентних та еферентних волокон було встановлено ще на початку минулого століття та отримало назву закону Белла – Мажанді. Після перерізання передніх корінців на одному боці спостерігається повне вимкнення рухових реакцій; Проте чутливість цієї сторони тіла зберігається. Перерізка задніх корінців вимикає чутливість, але з призводить до втрати рухових реакцій мускулатури.
1 – біла речовина;
2 – сіра речовина;
3 - задній (чутливий) корінець;
4 – спинно-мозкові нерви;
5 - передній (руховий) корінець;
6 - спинно-мозковий ганглій
Нейрони спинномозкових гангліїв відносяться до простих уніполярних, або псевдоуніполярних нейронів. Назва «псевдоуніполярний» пояснюється тим, що в ембріональному періоді первинні аферентні нейрони походять від біполярних клітин, відростки яких потім зливаються. Нейрони спинномозкових гангліїв можна поділити на клітини малих та великих розмірів. Тіло великих нейронів має діаметр близько 60-120 мкм, тоді як у дрібних нейронів він коливається від 14 до 30 мкм.
Великі нейрони дають початок товстим мієлінізованим волокнам. Від дрібних починаються як тонкі мієлінізовані, так і немієлінізовані волокна. Після біфуркації обидва відростки прямують у протилежні напрямки: центральний входить у дорсальний корінець і в його складі – у спинний мозок, периферичний – у різні соматичні та вісцеральні нерви, що підходять до рецепторних утворень шкіри, м'язів та внутрішніх органів.
Іноді центральні відростки первинних аферентних нейронів заходять до вентрального корінця. Це відбувається при трифуркації аксона первинного аферентного нейрона, внаслідок якої його відростки проектуються в спинний мозок і через дорсальний та вентральний коріння.
З усієї популяції клітин дорсальних гангліїв приблизно 60-70% належить до дрібних нейронів. Це відповідає тому, що число неміелінізованих волокон у дорсальному корінці перевищує число мієлінізованих волокон.
Тіла нейронів спинномозкових гангліїв немає дендритних відростків і отримують синоптичних входів. Їх збудження відбувається в результаті приходу потенціалу дії з периферичного відростка, що контактує з рецепторами.
Клітини спинномозкових гангліїв містять високі концентрації глутамінової кислоти – одного з передбачуваних медіаторів. Їх поверхнева мембрана містить рецептори, специфічно чутливі до g-аміномасляної кислоти, що збігається з високою чутливістю до g-аміномасляної кислоти центральних закінчень первинних аферентних волокон. Малі нейрони гангліїв містять речовину Р або соматостатин. Обидва ці поліпептиди також є ймовірними медіаторами, що вивільняються закінченнями первинних аферентних волокон.
Кожна пара корінців відповідає одному з хребців і залишає хребет через отвір між ними. Тому сегменти спинного мозку прийнято позначати за тим хребцем, біля якого із спинного мозку виходять відповідні корінці. Спинний мозок прийнято також розділяти на кілька відділів: шийний, грудний, поперековий і крижовий, кожен з яких містить кілька сегментів. У зв'язку з розвитком кінцівок нейронний апарат тих сегментів спинного мозку, які їх іннервують, набув найбільшого розвитку. Це знайшло своє відображення в утворенні шийного та поперекового потовщення. В області потовщення спинного мозку коріння містять найбільшу кількість волокон і мають найбільшу товщину.
На поперечному зрізі спинного мозку ясно виділяється центрально розташована сіра речовина, утворена скупченням нервових клітин, і біла речовина, що облямовує його, утворена нервовими волокнами. У сірій речовині розрізняють вентральні та дорсальні роги, між якими лежить проміжна зона. Крім того, у грудних сегментах розрізняють також бічне випинання сірої речовини – бічні роги.
Усі нейронні елементи спинного мозку можуть бути поділені на 4 основні групи: еферентні нейрони, вставні нейрони, нейрони висхідних трактів та інтраспінальні волокна чутливих аферентних нейронів. Моторні нейрони зосереджені в передніх рогах, де вони утворюють специфічні ядра, всі клітини яких посилають свої аксони до певного м'яза. Кожне рухове ядро зазвичай тягнеться кілька сегментів. Тому і аксони мотонейронів, що іннервують один і той же м'яз, залишають спинний мозок у складі декількох вентральних корінців.
Крім моторних ядер, розташованих у вентральних рогах, виділяються великі скупчення нервових клітин у проміжній зоні сірої речовини. Це основне ядро вставних нейронів спинного мозку. Аксони вставкових нейронів поширюються як усередині сегмента, і у найближчі сусідні сегменти.
Характерне скупчення нервових клітин займає також дорсальну частину дорсального роги. Ці клітини утворюють густі переплетення, а вказана зона отримала назву желатинозної субстанції Роланда.
Найбільш точне та систематизоване уявлення про топографію нервових клітин сірої речовини спинного мозку дає поділ його на послідовні шари, або пластини, у кожній з яких групуються головним чином однотипні нейрони.
Хоча пошарова друкарня сірої речовини була спочатку виявлена в спинному мозку кішки, вона виявилася досить універсальною і цілком застосовна до спинного мозку як інших хребетних, так і людини.
Згідно з цими даними, всю сіру речовину можна розділити на 10 пластин. Найперша дорсальна пластина містить головним чином так звані крайові нейрони. Їхні аксони проектуються рострально, даючи початок спиноталамічного тракту. На крайових нейронах закінчуються волокна тракту Ліссауера, який утворений сумішшю первинних аферентних волокон та аксонами пропріоспінальних нейронів.
Друга та третя пластини утворюють желатинозну субстанцію. Тут локалізуються два основних типи нейронів: дрібніші та відносно великі нейрони. Хоча тіла нейронів другої пластини мають невеликий діаметр, їх дендритні розгалуження дуже численні. Аксони нейронів другої пластини проектуються на тракт Ліссауера та власний дорсолатеральний пучок спинного мозку, але багато хто залишається в межах желатинозної субстанції. На клітинах другої та третьої пластин закінчуються волокна первинних аферентних нейронів, переважно шкірної та больової чутливості.
Четверта пластина займає центр дорсального рогу. Дендрит нейронів IV шару проникають в желатинозну субстанцію, а їх аксони проектуються в таламус і бічне цервікальне ядро. Синаптичні входи вони одержують від нейронів желатинозної субстанції, які аксони проектуються в таламус і бічне цервікальне ядро. Синаптичні входи вони одержують від нейронів желатинозної субстанції та первинних аферентних нейронів.
У цілому нині нервові клітини першої-четвертої пластин захоплюють всю вершину дорсального роги й утворюють первинну сенсорну область спинного мозку. Сюди проектуються волокна переважно дорсально-корінкових аферентів від екстерорецепторів, включаючи шкірну та больову чутливість. У цій же зоні локалізовані нервові клітини, що дають початок декільком висхідним трактам.
У п'ятій і шостій пластинах локалізуються численні типи вставних нейронів, які отримують синаптичні входи від волокон заднього корінця і низхідних шляхів, особливо кортико-спінального та руброспінального тракту.
У сьомій та восьмій пластинах локалізуються пропріоспінальні вставні нейрони, що дають початок довгим аксонам, що досягають нейронів віддалених сегментів. Тут закінчуються аферентні волокна від пропріорецепторів, волокна вестибулоспінального та ретикулоспінального трактів, аксони пропріоспінальних нейронів.
У дев'ятій пластині розташовуються тіла a- та g-мотонейронів. Цієї області досягають також пресинаптичні закінчення первинних афферентних волокон від м'язових рецепторів розтягування, закінчення волокон низхідних трактів, кортико-спінальних волокон, терміналі аксонів збуджуючих та гальмівних вставних нейронів.
Десята пластина оточує спинномозковий канал і містить поряд з нейронами значну кількість гліальних клітин та комісуральних волокон.
Клітини нейроглії спинного мозку на значному протязі покривають поверхню нейронів, причому відростки гліальної клітини спрямовані, з одного боку, до тіл нейронів, а з іншого, часто контактують з кровоносними капілярами, будучи посередниками між нервовими елементами та джерелами їх живлення.
Спинний мозок висхідними шляхами передає сигнали в надсегментарні рівні головного мозку, а по низхідних отримує звідти команди до дії. Східні шляхи передають імпульси від пропріоцепторів по волокнах спинобульбарних пучків Голля і Бурдаха і спинномозочкових шляхів Говерса і Флексіго, від больових і температурних рецепторів по латеральному спиноталамічному тракту, від тактильних рецепторів по вентральному спиноталамічному шляху.
Східні шляхи проходять у складі кортикоспінальних, або пірамідних, трактів та екстракортикоспінальних, або екстрапірамідних.
Для контролю над роботою внутрішніх органів, рухових функцій, своєчасного отримання та передачі симпатичних та рефлекторних імпульсів, використовуються провідні шляхи спинного мозку. Порушення передачі імпульсів призводить до серйозних збоїв у роботі всього організму.
У чому полягає провідна функція спинного мозку
Під терміном «провідні шляхи» мається на увазі сукупність нервових волокон, що забезпечують передачу сигналів у різні центри сірої речовини. Висхідні та низхідні шляхи спинного мозку виконують основну функцію – передачу імпульсів. Прийнято розрізняти три групи нервових волокон:- Асоціативні провідні шляхи.
- Комісуральні зв'язки.
- Проекційні нервові волокна.
Чутливі та рухові шляхи забезпечують міцний взаємозв'язок між спинним та головним мозком, внутрішніми органами, м'язовою системоюта опорно-руховим апаратом. Завдяки швидкій передачі імпульсів, всі рухи тіла здійснюються злагодженим чином, без відчутних зусиль з боку людини.
Чим утворені провідні спинномозкові шляхи
Основні провідні шляхи утворені зв'язками клітин – нейронів. Така будова забезпечує необхідну швидкість передачі імпульсів.Класифікація провідних шляхів залежить від функціональних особливостей нервових волокон:
- Висхідні провідні шляхи спинного мозку – зчитують і передають сигнали: зі шкіри та слизових оболонок людини, органів життєзабезпечення. Забезпечують виконання функцій опорно-рухового апарату.
- Східні провідні шляхи спинного мозку – передають імпульси безпосередньо робочим органам тіла людини – м'язовим тканинам, залозам тощо. З'єднані безпосередньо з кірковою частиною сірої речовини. Передача імпульсів відбувається через спинномозковий нейронний зв'язок, до внутрішніх органів.
Спинний мозок має подвійний напрямок провідних шляхів, що забезпечує швидку імпульсну передачу інформації від контрольованих органів. Провідникова функція спинного мозку здійснюється завдяки наявності ефективної передачі імпульсів нервової тканини.
У медичній та анатомічній практиці прийнято використовувати такі терміни: 
Де розташовані провідні шляхи мозку спини
Всі нервові тканини розташовуються в сірій та білій речовині, з'єднують спинномозкові роги та кору півкуль.Морфофункціональна характеристика низхідних провідних шляхів спинного мозку обмежує напрямок імпульсів лише в одному напрямку. Роздратування синапсів здійснюється від пресинаптичної до постсинаптичної мембрани.
Провідникової функції спинного та головного мозку відповідають наступні можливості та розташування основних висхідних та поблажливих шляхів:
- Асоціативні провідні шляхи є «містками», що з'єднують ділянки між корою та ядрами сірої речовини. Складаються з коротких та довгих волокон. Перші, що знаходяться в межах однієї половини або частки мозкових півкуль.
Довгі волокна здатні передавати сигнали через 2-3 сегменти сірої речовини. У спинномозковій речовині нейрони утворюють міжсегментарні пучки. - Комісуральні волокна - утворюють мозолисте тіло, що сполучає новостворені відділи спинного та головного мозку. Розходяться променистим способом. Розташовані у білій речовині мозкової тканини.
- Проекційні волокна – місце розташування провідних шляхів у спинному мозку дозволяє імпульсам максимально швидко досягати кори півкуль. За характером та функціональними особливостями, проекційні волокна поділяються на висхідні (аферентні шляхи) та низхідні.
Перші поділяють на екстерорецептивні (зір, слух), пропріорецептивні (рухові функції), інтерорецептивні (зв'язок із внутрішніми органами). Рецептори розташовуються між хребтовим стовпом та гіпоталамусом.

Анатомія провідних шляхів досить складна для людини, яка не має медичної освіти. Але нейронна передача імпульсів є тим, що робить організм людини єдиним цілим.
Наслідки пошкоджень провідних шляхів
Щоб зрозуміти нейрофізіологію сенсорних та рухових шляхів, слід трохи познайомитися з анатомією хребта. Спинний мозок має структуру, яка багато в чому нагадує циліндр, оточений м'язовою тканиною. Усередині сірої речовини проходять провідні шляхи, що контролюють роботу внутрішніх органів, а також рухові функції. Асоціативні провідні шляхи відповідають за болючі та тактильні відчуття. Двигуни – за рефлекторні функції організму.
Усередині сірої речовини проходять провідні шляхи, що контролюють роботу внутрішніх органів, а також рухові функції. Асоціативні провідні шляхи відповідають за болючі та тактильні відчуття. Двигуни – за рефлекторні функції організму.
Внаслідок травми, вад розвитку або захворювань спинного мозку, провідність може знизитися або повністю припинитися. Відбувається це через відмирання нервових волокон. Для повного порушення провідності імпульсів спинного мозку характерна паралізація, відсутність чутливості кінцівок. Починаються збої в роботі внутрішніх органів, за які відповідає пошкоджений нейронний зв'язок. Так, при ураженні нижньої частини спинного мозку, спостерігається нетримання сечі та мимовільна дефекація.
Рефлекторна та провідникова діяльність спинного мозку порушується одразу після виникнення дегенеративних патологічних змін. Відбувається відмирання нервових волокон, які важко піддаються відновленню. Хвороба швидко прогресує та настає грубе порушення провідності. З цієї причини приступати до медикаментозному лікуваннюнеобхідно якомога раніше.
Як відновити прохідність у спинному мозку
Лікування непровідності насамперед пов'язане з необхідністю припинення відмирання нервових волокон, а також усунення причин, що стали каталізатором патологічних змін.Медикаментозне лікування
Полягає у призначенні препаратів, що перешкоджають відмирання клітин мозку, а також достатньому кровопостачанню пошкодженої ділянки спинного мозку. При цьому враховуються вікові особливості функції спинного мозку, що проводить, а також серйозність травми або захворювання.Для додаткової стимуляції нервових клітин проводиться лікування електричними імпульсами, що допомагає підтримувати тонус м'язів.
Хірургічне лікування
Операція з відновлення провідності спинного мозку торкається двох основних напрямків:- Усунення каталізаторів, що спричинили паралізацію роботи нейронних зв'язків.
- Стимуляція спинного мозку для відновлення втрачених функцій.
Народна медицина при порушенні провідності
Народні засоби при порушенні провідності спинного мозку, якщо й використовуються, повинні застосовуватися особливо обережно, щоб не призвести до погіршення стану пацієнта.Особливою популярністю користуються: 
Повністю відновити нейронні зв'язки після травм досить складно. Багато що залежить від швидкого звернення до медичний центрта кваліфікованої допомоги нейрохірурга. Чим більше часу мине від початку дегенеративних змін, тим менше шансів на відновлення функціональних можливостей спинного мозку.
Морфофункціональна організація спинного мозку
Спинний мозок – найдавніший відділ ЦНС хребетних. Він уже є у ланцетника, найпримітивнішого представника хордових.
Спинний мозок – каудальний відділ центральної нервової системи. Він міститься в хребетному каналі і має неоднакову довжину у різних представників хребетних.
У людини в каутальному відділі хребетного каналу збираються коріння каудальних відділів спинного мозку, утворюючи так званий кінський хвіст.
Спинний мозокхарактеризується сегментарною будовою. У спинному мозку виділяють шийний, грудний, поперековий, крижовий та куприковий відділи. Кожен відділ складається з кількох сегментів. Шийний відділ включає 8 сегментів (З 1 – З 8), грудний – 12 (Th 1 – Th 12), поперековий – 5 (L 1 – L 5), крижовий – 5 (S 1 – S 5) та куприковий – 1- 3 (З 1 - Co 3). Від кожного сегмента відходять дві пари корінців, які відповідають одному з хребців і покидають хребет через отвір між ними.
Розрізняють дорсальні (задні) та вентральні (передні) коріння. Дорсальні коріння утворені центральними аксонами первинних аферентних нейронів, тіла яких лежать у спинномозкових гангліях.
Вентральні коріння сформовані аксонами α- та γ-мотонейронів та немієлінізованими волокнами нейронів вегетативної нервової системи. Такий розподіл аферентних та еферентних волокон було встановлено незалежно один від одного на початку XIX століття Ч. Беллом (1811) та Ф. Мажанді (1822). Різне розподіл функцій у передніх і задніх корінцях спинного мозку отримало назву закону Белла-Мажанді. Сегменти спинного мозку та хребці відповідають одному метамеру. Нервові волокна пари задніх корінців йдуть як до свого метамеру, але й вище й нижче – у сусідні метамери. Шкірна область, де розподіляються ці чутливі волокна, називається дерматомом.
Кількість волокон у дорсальному корінці набагато більша, ніж у вентральному.
Нейронні структури спинного мозку.Центральну частину поперечного зрізу спинного мозку займає сіра речовина. Навколо сірої речовини розташовується біла речовина. У сірій речовині виділяють передні, задні та бічні роги, а в білому – стовпи (вентральні, дорсальні, латеральні тощо).
Нейрональний склад спинного мозку досить різноманітний. Виділяють кілька видів нейронів. Тіла нейронів спинномозкових гангліїв знаходяться поза спинним мозком. У спинний мозок входять аксони цих нейронів. Нейрони спинномозкових гангліїв – це уніполярні або псевдоуніполярні нейрони. У спинномозкових гангліях лежать тіла соматичних аферентів, що іннервують в основному скелетні м'язи. Тіла інших чутливих нейронів знаходяться в тканині та в інтрамуральних гангліях автономної нервової системи та забезпечують чутливість лише внутрішніх органів. Вони бувають двох типів: великі – діаметром 60-120 мкм та дрібні – діаметром 14-30 мкм. Великі дають мієлінізовані волокна, а дрібні – мієлінізовані та неміелінізовані. Нервові волокна чутливих клітин за швидкістю проведення та діаметром класифікуються на А-, В- та С-волокна. Товсті мієлінізовані А-волокнадіаметром від 3 до 22 мкм та швидкістю проведення від 12 до 120 м/с поділяються на підгрупи: альфа-волокна – від м'язових рецепторів, бета-волокна – від тактильних та барорецепторів, дельта-волокна – від терморецепторів, механорецепторів та больових рецепторів. До волокнам групи Ввідносять мієлінізовані волокна середньої товщини зі швидкістю проведення збудження 3-14 м/с. По них переважно передається відчуття болю. До аферентам типу Свідносять більшість безмієлінових волокон товщиною не більше 2 мкм та швидкістю проведення до 2 м/с. Це волокна, що йдуть від больових, хемо- та деяких механорецепторів.
У сірій речовині спинного мозку виділяють такі елементи:
1) еферентні нейрони (мотонейрони);
2) вставні нейрони;
3) нейрони висхідних трактів;
4) інтраспінальні волокна чутливих аферентних нейронів.
Моторні нейронизосереджені у передніх рогах, де вони утворюють специфічні ядра, всі клітини яких посилають свої аксони до певного м'яза. Кожне рухове ядро тягнеться зазвичай протягом декількох сегментів, тому і їх аксони, які іннервують один і той самий м'яз, залишають спинний мозок у складі декількох вентральних корінців.
У проміжній зоні сірої речовини локалізуються вставні нейрони. Їх аксони поширюються як усередині сегмента, і у найближчі сусідні сегменти. Вставні нейрони– різнорідна група, дендрити та аксони якої не залишають меж спинного мозку. Вставні нейрони утворюють синаптичні контакти тільки з іншими нейронами та їх більшість. Перед вставкових нейронів припадає близько 97% від усіх нейронів. За розмірами вони менші за α-мотонейрони, здатні до високочастотної імпульсації (вище 1000 в сек.). Для пропріоспінальних вставних нейронівХарактерна властивість посилати довгі аксони через кілька сегментів і закінчуватися на мотонейронах. Разом про те ці клітини конвергують волокна різних низхідних трактів. Тому вони є релейними станціями на шляху від нейронів до мотонейронів. Особливу групу вставних нейронів утворюють гальмівні нейрони. До них відносяться, наприклад, клітки Реншоу.
Нейрони висхідних трактівтакож повністю перебувають у межах ЦНС. Тіла цих нейронів розташовані у сірій речовині спинного мозку.
Центральні закінчення первинних аферентівмають свої особливості. Після вступу в спинний мозок аферентне волокно, як правило, дає початок висхідній та низхідній гілкам, які можуть йти на значні відстані по спинному мозку. Кінцеві розгалуження одного нервового аферентного волокна мають численні синапси одному мотонейроні. Крім того, встановлено, що одне волокно, що йде від рецептора розтягування, утворює синапс майже з усіма мотонейронами даного м'яза.
У дорсальній частині дорсального рогу міститься желатинозна субстанція Роланда.
Найточніше уявлення про топографію нервових клітин сірої речовини спинного мозку дає поділ його на послідовні шари або пластини, у кожній з яких групуються, як правило, однотипні нейрони.
Згідно з цими даними, вся сіра речовина спинного мозку була поділена на 10 пластин (Рексед) (рис. 2.2).
I – крайові нейрони – дають початок спиноталамічного тракту;
II-III – желатинозна субстанція;
I-IV – загалом первинна сенсорна область спинного мозку (аферентація від екстерорецепторів, аферентація від рецепторів шкірної та больової чутливості);
Рис. 2.2.Розподіл сірої речовини спинного мозку на пластини (за Рекседом)
V-VI – локалізуються вставні нейрони, які отримують входи від задніх корінців та низхідних шляхів (кортико-спінальний, рубро-спінальний);
VII-VIII – розташовуються пропріоспінальні вставкові нейрони (від пропріорецепторів, волокон вестибуло-спінального та ретикуло-спі-
ного трактів), аксони пропріоспінальних нейронів;
IX – містить тіла α- та γ-мотонейронів, пресинаптичні волокна первинних аферентів від м'язових рецепторів розтягування, закінчення волокон низхідних трактів;
X - оточує спинномозковий канал і містить поряд з нейронами значну кількість гліальних клітин та комісуральних волокон.
Властивості нервових елементів спинного мозку.Спинний мозок людини містить приблизно 13 мільйонів нейронів.
α-мотонейрони – великі клітини з довгими дендритами, що мають до 20 000 синапсів, більша частина яких утворена закінченнями інтраспінальних вставних нейронів. Швидкість проведення їх аксону становить 70-120 м/с. Характерні ритмічні розряди із частотою не вище 10-20 імп/с, що пов'язано з вираженою слідовою гіперполяризацією. Це вихідні нейрони. Вони здійснюють передачу сигналів скелетним м'язовим волокнам, вироблених у спинному мозку.
γ-мотонейрони – дрібніші клітини. Діаметр їх трохи більше 30-40 мкм, вони мають безпосереднього контакту з первинними аферентами.
γ-мотонейрони іннервують інтрафузальні (внутрішньоверетенні) м'язові волокна.
Вони моносинаптично активуються волокнами низхідних трактів, що відіграє важливу роль в α-, γ-взаємодії. Швидкість проведення за аксоном нижче – 10-40 м/с. Частота імпульсів вище, ніж у α-мото-
нейронів – 300-500 імп/с.
У бічних і передніх рогах знаходяться прегангліонарні нейрони вегетативної нервової системи – аксони їх спрямовуються до клітин гангліїв симпатичного нервового ланцюжка та інтрамуральних ганглій внутрішніх органів.
Тіла симпатичних нейронів, аксони яких утворюють преганглионарные волокна, розташовуються в интермедиолатеральном ядрі спинного мозку. Їхні аксони відносяться до групи В-волокон. Їм властива стала тонічна імпульсація. Одні з цих волокон беруть участь у підтримці судинного тонусу, інші забезпечують регуляцію вісцеральних эффекторных структур (гладкої мускулатури травної системи, залізистих клітин).
Тіла парасимпатичних нейронів утворюють крижові парасимпатичні ядра. Вони розташовуються у сірій речовині крижового відділу спинного мозку. Для багатьох їх характерна фонова імпульсна активність, частота якої зростає, наприклад, у міру підвищення тиску в сечовому міхурі.
Спинний мозок і двох симетричних половин, відмежованих друг від друга спереду - глибокої серединної щілиною, а ззаду - серединної борозеною. Спинний мозок характеризується сегментарною будовою; з кожним сегментом пов'язана пара передніх (вентральних) та пара задніх (дорсальних) корінців.
У спинному мозку розрізняють сіру речовину, розташовану в центральній частині, та білу речовину, що лежить по периферії.
Біла речовина спинного мозку є сукупністю поздовжньо орієнтованих переважно мієлінових нервових волокон. Пучки нервових волокон, що здійснюють зв'язок між різними відділами нервової системи, називаються трактами або провідними шляхами спинного мозку.
Сіра речовина на поперечному розрізі має вигляд метелика і включає передні, або вентральні, задні, або дорсальні, і бічні, або латеральні роги. У сірій речовині знаходяться тіла, дендрити та (частково) аксони нейронів, а також гліальні клітини. Основний складовоюсірої речовини є мультиполярні нейрони.
Клітини, подібні за розмірами, тонкою будовою та функціональним значенням, лежать у сірій речовині групами, які називаються ядрами.
Аксони корінцевих клітин залишають спинний мозок у складі його передніх корінців. Відростки внутрішніх клітин закінчуються синапсами у межах сірої речовини спинного мозку. Аксони пучкових клітин проходять у білій речовині відокремленими пучками волокон, що несуть нервові імпульси від певних ядер спинного мозку до інших сегментів або у відповідні відділи головного мозку, утворюючи провідні шляхи. Окремі ділянки сірої речовини спинного мозку значно відрізняються одна від одної за складом нейронів, нервових волокон та нейроглії.
У задніх рогах розрізняють губчастий шар, желатинозну речовину, власне ядро заднього рогу та грудне ядро Кларка. Між задніми і бічними рогами сіра речовина вдається тяжами в біле, внаслідок чого утворюється його сіткоподібне розпушення, що отримало назву сітківки, або ретикулярної формації, спинного мозку.
Задні роги багаті дифузно розташованими вставними клітинами. Це дрібні мультиполярні асоціативні та комісуральні клітини, аксони яких закінчуються в межах сірої речовини спинного мозку тієї ж сторони (асоціативні клітини) або протилежної сторони (комісуральні клітини).
Нейрони губчастої зони та желатинозної речовини здійснюють зв'язок між чутливими клітинами спинальних гангліїв та руховими клітинами передніх рогів, замикаючи місцеві рефлекторні дуги.
Нейрони ядра Кларка отримують інформацію від рецепторів м'язів, сухожиль і суглобів (пропріоцептивна чутливість) по товстим корінцевим волокнам і передають її в мозок.
У проміжній зоні розташовані центри вегетативної (автономної) нервової системи – прегангліонарні холінергічні нейрони її симпатичного та парасимпатичного відділів.
У передніх рогах розташовані найбільші нейрони спинного мозку, які утворюють значні за обсягом ядра. Це так само, як і нейрони ядер бічних рогів, корінцеві клітини, оскільки їх нейрит складають основну масу волокон передніх корінців. У складі змішаних спинномозкових нервів вони надходять на периферію та утворюють моторні закінчення у скелетній мускулатурі. Таким чином, ядра передніх рогів є моторними соматичними центрами.
Нервову систему прийнято поділяти на кілька відділів. За топографічними ознаками її ділять на центральний та периферичний відділи, за функціональними ознаками – на соматичний та вегетативний відділи. Центральний відділ, або центральна нервова система, включає головний та спинний мозок. До периферичного відділу, або периферичної нервової системи, відносять усі нерви, тобто всі периферичні провідні шляхи, які складаються з чутливих та рухових нервових волокон. Соматичний відділ, або соматична нервова система, включає черепномозкові та спинномозкові нерви, що пов'язують центральну нервову систему з органами, що сприймають зовнішні подразнення - зі шкірним покривом та апаратом руху. Вегетативний відділ, або вегетативна нервова система, забезпечує зв'язок центральної нервової системи з усіма внутрішніми органами, залозами, судинами та органами, у складі яких є гладка м'язова тканина. Вегетативний відділ ділиться на симпатичну та парасимпатичну частини, або симпатичну та парасимпатичну нервову систему.
До складу центральної нервової системи входять головний та спинний мозок. Між масою головного та спинного мозку є певні співвідношення: у міру підвищення організації тварини збільшується відносна масаголовного мозку порівняно зі спинним. У птахів головний мозок у 1,5-2,5 рази більше спинного, у копитних – у 2,5-3, у хижих – у 3,5-5, у приматів – у 8-15 разів.
Спинний мозок- medulla spinalis лежить у хребетному каналі, займаючи приблизно 2/3 його обсягу. У великої рогатої худоби та коня його довжина дорівнює 1,8-2,3 м, маса 250-300 г, у свині - 45-70 г. Він має вигляд циліндричного тяжа, дещо сплюснутого дорсовентрально. Чіткої межі між головним та спинним мозком немає. Вважається, що вона відбувається на рівні краніального краю атланту. У спинному мозку розрізняють шийну, грудну, поперекову, крижову та хвостову частини за місцем їх залягання. У ембріональний період розвитку спинний мозок заповнює весь хребетний канал, але у зв'язку з великою швидкістю зростання скелета різниця в їхній довжині стає дедалі більшою. В результаті мозок у великої рогатої худоби закінчується на рівні 4-го, у свині – в області 6-го поперекового хребця, а у коня – в області 1-го сегмента крижової кістки. Уздовж спинного мозку з його дорсальної стороні проходить серединна дорсальна борозна (жолоб). Від неї вглиб відходить сполучнотканинна дорсальна перегородка. З боків або серединної борозни йдуть дрібніші дорсальні латеральні борозни. По вентральній стороні йде глибока серединна вентральна щілина, а з боків від неї – вентральні латеральні борозни (жолоби). Наприкінці спинний мозок різко звужується, утворюючи мозковий конус, який переходить у кінцеву нитку. Вона утворена сполучною тканиною і закінчується лише на рівні перших хвостових хребців.
У шийній та поперековій частинах спинного мозку є потовщення. У зв'язку з розвитком кінцівок у цих ділянках збільшується кількість нейронів та нервових волокон. У свині шийне потовщення сформоване 5-8 нейросегментами. Його максимальна ширина лише на рівні середини 6-го шийного хребця дорівнює 10 мм. Поперекове потовщення припадає на 5-7-й поперекові нейросегменти. У кожному сегменті від спинного мозку відходить двома корінцями пара спинномозкових нервів – праворуч та зліва. Дорсальний корінець відходить від дорсальної латеральної борозни, вентральний корінець - від вентральної латеральної борозни. З хребетного каналу спинномозкові нерви виходять через міжхребцеві отвори. Ділянка спинного мозку між двома сусідніми спинномозковими нервами називається нейросегментом. Нейросегменти бувають різної довжини та часто за розмірами не відповідають довжині кісткового сегмента. В результаті спинномозкові нерви відходять під різним кутом. Багато хто з них проходить деяку відстань усередині хребетного каналу до виходу з міжхребцевого отвору свого сегмента. У каудальному напрямку ця відстань збільшується і з нервів, що йдуть усередині хребетного каналу, за мозковим конусом утворюється ніби пензлик, названий «кінським хвостом».
Головний мозок- encephalon - міститься в черепномозковій коробці і складається з нескільки частин. У копитних відносна маса головного мозку 0,08-0,3% від маси тіла, що становить у коня 370-600 г, у великої рогатої худоби - 220-450, у вівці та свині - 96-150 г. У дрібних тварин відносна маса головного мозку зазвичай більша, ніж у великих.
Головний мозок копитних напівовальної форми. У жуйних - з широкою фронтальною площиною, з нюховими цибулинами, що майже не виступають, і помітними розширеннями на рівні скроневих областей. У свині - більш звужений попереду, з нюховими цибулинами, що помітно виступають. Довжина його складає в середньому у великої рогатої худоби 15 см, у вівці - 10, у свині-11 см. Глибокою поперечною щілиною головний, мозок ділиться на великий мозок, що лежить рострально, і ромбоподібний мозок, розташований каудальний. Ділянки головного мозку філогенетично древні, що є продовження проекційних провідних шляхів спинного мозку, називаються стовбуром мозку. Він включає довгастий мозок, мозковий міст, середній міст, частина проміжного мозку. Філогенетично молодші частини головного мозку утворюють покривну частину мозку. У неї входять півкулі головного мозку та мозочок.
Ромбоподібний мозок- rhombencephalon - ділиться на довгастий та задній мозок і містить четвертий мозковий шлуночок.
Продовгуватий мозок- medulla oblongata - найзадніша ділянка головного мозку. Його маса становить 10-11% маси мозку; довжина у великої рогатої худоби - 4,5, у вівці - 3,7, у свині - 2 див. .
На його дорсальній стороні є поглиблення ромбовидної форми – четвертий мозковий шлуночок. По вентральній стороні проходять три борозни: серединна та 2 бічні. З'єднуючись каудально, вони переходять у вентральну серединну щілину спинного мозку. Між борознами лежать 2 вузькі подовжені валики - піраміди, в яких проходять пучки рухових нервових волокон. На межі довгастого та спинного мозку пірамідні тракти перехрещуються – утворюється перехрест пірамід. У довгому мозку сіра речовина розташована всередині, в дні четвертого мозкового шлуночка у вигляді ядер, що дають початок черепномозковим нервам (з VI по XII пару), а також ядер, в яких відбувається перемикання імпульсів на інші відділи головного мозку. Біла речовина лежить зовні, переважно вентрально, формуючи провідні шляхи. Двигуни (еферентні) провідні шляхи з головного мозку у спинній формують піраміди. Чутливі провідні шляхи (аферентні) зі спинного мозку в головний утворюють/задні ніжки мозочка, що йдуть з довгастого мозку до мозочка. У масі довгастого мозку як сітчастого сплетення залягає важливий координаційний апарат мозку - ретикулярна формація. Вона поєднує структури стовбура мозку і сприяє їх залученню в складні, багатоступінчасті реакції у відповідь.
Продовгуватий мозок- життєво важлива ділянка центральної нервової системи (ЦНС), її руйнування призводить до миттєвої смерті. Тут розташовані центри дихання, серцебиття, жування, ковтання, ссання, блювання, жуйки, слино- та соковиділення, тонусу судин та ін.
Задній мозок- Metencephalon - складається з мозочка і мозкового мосту.
Мозковий міст- pons - масивне потовщення на вентральній поверхні мозку, що лежить поперек передньої частини довгастого мозку шириною до 3,5 см у великої рогатої худоби, 2,5 см у вівці і 1,8 ом у свині. Основну масу мозкового мосту складають провідні шляхи (низхідні та висхідні), що з'єднують головний мозок зі спинним та окремі ділянки головного мозку між собою. Велика кількістьнервових волокон йде поперек моста до мозочка і формує середні ніжки мозочка. У мосту розташовані групи ядер, зокрема ядра черепномозкових нервів (V пара). Від бічної поверхні мосту відходить найбільша V пара черепномозкових нервів – трійчасті.
Мозочок- cerebellum - розташовується над мостом, довгастим мозком і четвертим мозковим шлуночком, позаду четверохолмія. Спереду межує із півкулями великого мозку. Маса його становить 10–11% маси мозку. У вівці та свині довжина його (4-4,5 см) більша за висоту (2,2-2,7 ом), у великої рогатої худоби наближається до кулястої - 5,6X6,4 см. У мозочку розрізняють середню частину - черв'ячок і бічні частини - півкулі мозочка. Мозочок має 3 пари ніжок. Задніми ніжками (мотузковими тілами) він з'єднаний з довгим мозком, середніми з мозковим мостом, передніми (ростральними) - із середнім мозком. Поверхня мозочка зібрана в численні складчасті часточки і звивини, розділені борознами та щілинами. Сіра речовина в мозочку розташована зверху - кора мозочка і в глибині у вигляді ядер. Поверхня кори мозочка у великої рогатої худоби становить 130 см 2 (близько 30% до кори великих півкуль) при товщині 450-700 мкм. Біла речовина розташована під корою та має вигляд гілки дерева, за що названо древом життя.
Мозочок є центром координації довільних рухів, підтримки тонусу м'язів, пози, рівноваги.
Ромбоподібний мозокмістить четвертий мозковий шлуночок. Його дном є поглиблення довгастого мозку – ромбоподібна ямка. Його стінки утворені ніжками мозочка, а дах переднім (ростральним) та заднім мозковими вітрилами, які є судинним сплетенням. Шлуночок повідомляється рострально з мозковим водопроводом, каудально – з центральним каналом спинного мозку та через отвори в вітрилі – з підпаутинним простором.
Великий мозок- cerebrum - включає кінцевий, проміжний і середній мозок. Кінцевий та проміжний мозок об'єднані в передній мозок.
Середній мозок - mesencephalon - складається з четверохолмія, ніжок великого мозку та укладеного між ними мозкового водопроводу. Прикритий великими півкулями. Його маса становить 5-6% від маси мозку.
Четверохолмія утворює дах середнього мозку. Воно складається з пари ростральних (передніх) горбків та пари каудальних (задніх) горбків. Четверохолмія є центром безумовно-рефлекторних рухових актів у відповідь на зорові та слухові роздратування. Передні пагорби вважаються підкірковими центрами зорового аналізатора, задні пагорби-підкірковими центрами слухового аналізатора. У жуйних передні пагорби більші за задні, у свині - навпаки.
Ніжки великого мозку утворюють дно середнього мозку. Мають вигляд двох товстих валиків, що лежать між зоровими трактами та мозковим мостом. Розділені міжніжковою борозеною.
Між чотиригорбом і ніжками великого мозку у вигляді вузької трубки проходить мозковий (сильвієвий) водогін. Рострально він з'єднується з третім, каудально – з четвертим мозковими шлуночками. Мозковий водогін оточений речовиною ретикулярної формації.
У середньому мозку біла речовина розташована зовні і являє собою провідні аферентні та еферентні шляхи. Сіра речовина розташована у глибині у вигляді ядер. Від мозкових ніжок відходить III пара черепномозкових нервів.
Проміжний мозок- diencephalon - складається з зорових пагорбів - таламуса, надгір'я - епіталамуса, підгір'я - гіпоталамуса. Розташований проміжний мозок між кінцевим.
У середнім мозком, прикритий кінцевим мозком. Його маса становить 8-9% від маси мозку. Зорові пагорби – найбільш масивна, центрально розташована частина проміжного мозку. Зростаючись між собою, вони здавлюють третій мозковий шлуночок так, що він набуває форми кільця, що йде навколо проміжної маси зорових пагорбів. Зверху шлуночок прикритий судинною покришкою; повідомляється міжшлуночковим отвором із бічними шлуночками, аборально переходить у мозковий водогін. Біла речовина в таламус лежить зверху, сіра - всередині у вигляді численних ядер. Вони служать перемикачами з нижніх відділів на кору і пов'язані майже з усіма аналізаторами. На базальній поверхні проміжного мозку розташований перехрест зорових нервів – хіазму.
Епітамус складається з декількох структур, у тому числі епіфіза та судинної покришки третього мозкового шлуночка (епіфіз – заліза внутрішньої секреції). Розташований у поглибленні між зоровими пагорбами та чотиригорбом.
Гіпоталамус розташований на базальній поверхні проміжного мозку між хіазмою та ніжками мозку. Складається з кількох частин. Безпосередньо за хіазмою у вигляді овального горбка - сірий бугор. Його звернена вниз верхівка витягнута за рахунок випинання стінки третього шлуночка і утворює вирву, на якій підвішено гіпофіз - залізо внутрішньої секреції. Позаду сірого бугра невелике округле утворення - соскоподібне тіло. Біла речовина в гіпоталамусі розташована зовні, формує провідні аферентні та еферентні шляхи. Сіра речовина - у вигляді численних ядер, оскільки гіпоталамус є найвищим підкірковим вегетативним центром. Він містить центри дихання, крово- та лімфообігу, температури, статевих функцій та ін.
Кінцевий мозок – telencephalon – утворений двома півкулями, розділеними глибокою поздовжньою щілиною та з'єднаними мозолистим тілом. Його маса у (великої рогатої худоби 250-300 г, у вівці та свині 60-80 г, що становить 62-66% від маси головного мозку. У кожній півкулі розрізняють дор-солатерально розташований плащ, вентромедіально - нюховий мозок, у глибині - смугасте тіло і бічний шлуночок Бакові шлуночки розділені прозорою перегородкою, з третім мозковим шлуночком повідомляються міжшлуночковим отвором.
Нюховий мозок складається з декількох частин, помітних на вентральній поверхні кінцевого мозку. Рострально, дещо виступаючи за межі плаща, лежать 2 нюхові цибулини. Вони займають ямки гратчастої кістки. Через отвір у продірявленій платівці кістки у них вступають нюхові нитки, які у сумі утворюють нюховий нерв. Цибулини є первинними нюховими центрами. Від них відходять нюхові тракти - аферентні провідні шляхи. Латеральний нюховий тракт доходить до грушовидних часток, розташованих латерально від ніжок мозку. Медіальні нюхові тракти досягають медіальної поверхні плаща. Між трактами лежать нюхові трикутники. Грушоподібні частки та нюхові трикутники – це вторинні нюхові центри. У глибині нюхового мозку, на дні бічних шлуночків, розташовані решта нюхового мозку. Вони пов'язують нюховий мозок коїться з іншими відділами мозку. Смугасте тіло розташоване в глибині півкуль і є базальним комплексом ядер, що є підкірковими руховими центрами.
Плащ досягає найбільшого розвитку у вищих ссавців. У ньому знаходяться найвищі центри всієї життєдіяльності тварини. Поверхня плаща вкрита звивинами та борознами. У великої рогатої худоби її поверхня дорівнює 600 см 2 . Сіра речовина в плащі розташована зверху – це кора великих півкуль. Біла речовина знаходиться всередині - це провідні шляхи. Функції різних ділянок кори нерівнозначні, будова відрізняється мозаїчністю, що дало можливість виділити в півкулях кілька часток (лобову, тім'яну, скроневу, потиличну) і кілька десятків полів. Поля відрізняються один від одного своєю цитоархітектонікою – розташуванням, кількістю та формою клітин та мієлоархітектонікою – розташуванням, кількістю та формою волокон.
Оболонки мозку -meninges. Спинний та головний мозок одягають тверда, павутинна та м'яка оболонки.
Тверда оболонка - поверхнева, товста, утворена щільною сполучною тканиною, бідна судинами. З кістками черепа та хребцями зростається зв'язками, складками та іншими утвореннями. Вона опускається в поздовжню щілину між півкулями великого мозку про вид серповидної зв'язки (серп великого мозку) і відокремлює великий мозок від ромбовидної перетинчастої палатки мозочка. Між нею та кістками є не скрізь розвинений епідуральний простір, заповнений пухкою сполучною та жировою тканинами. Тут проходять вени. Зсередини тверда мозкова оболонка вистелена зізнавством. Між нею та павутинною оболонкою є субдуральмий простір, заповнений спинномозковою рідиною. Павутинна оболонка - утворена пухкою сполучною тканиною, ніжна, безсудинна, борозни не заходить. З обох боків покрита ендотелією і відокремлена субдуральньф та яубарахноїдальним (підпаутинним) просторами від інших оболонок. Приєднується до оболонок за допомогою зв'язок, а також судин та нервів, що проходять через неї.
М'яка оболонка – тонка, але щільна, з великою кількістю судин, за що її називають також судинною. Заходить у всі борозни та щілини головного та спинного мозку, а також у мозкові шлуночки, де формує судинні покришки.
Міжболочкові простори, мозкові шлуночки та центральний спинномозковий канал заповнені спинномозковою рідиною, яка є внутрішнім середовищеммозку і захищає його від шкідливих впливів, регулює внутрішньочерепний тиск, виконує захисну функцію. Утворюється рідина. В основному в судинних покришках шлуночків, що відтікає у венозне русло. У нормі її кількість постійно.
Судини головного та спинного мозку. Спинний мозок постачається кров'ю по гілках, що відходять від хребетних, межребергаих, поперекових та крижових артерій. У хребетному каналі вони формують спинномозкові артерії, що йдуть у борозенах та центральній щілині спинного мозку. До головного мозку кров підходить за хребетними та внутрішніми сонними (у рогатої худоби - за внутрішніми щелепними) артеріями.