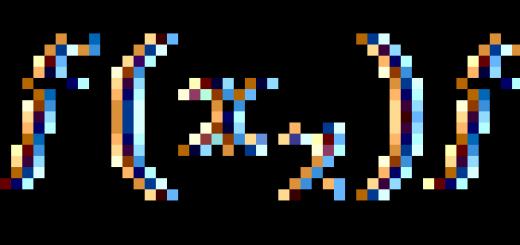Represents flattened strand located in the spinal canal, about 45 cm long in men and 42 cm in women. In places where the nerves exit to the upper and lower extremities, the spinal cord has two thickenings: cervical and lumbar.
The spinal cord is made up of two types of fabric: outer white (bundles of nerve fibers) and inner gray matter (nerve cell bodies, dendrites and synapses). In the center of the gray matter, a narrow channel with cerebrospinal fluid runs along the entire brain. The spinal cord has segmental structure(31-33 segments), each of its sections is associated with a specific part of the body, 31 pairs of spinal cords depart from the segments of the spinal cord nerves: 8 pairs of cervical (Ci-Cviii), 12 pairs of thoracic (Thi-Thxii), 5 pairs of lumbar (Li-Lv), 5 pairs of sacral (Si-Sv) and a pair of coccygeal (Coi-Coiii).
Each nerve divides into front and back roots. back roots- afferent pathways front roots efferent pathways. Afferent impulses from the skin, motor apparatus, and internal organs enter the spinal cord along the posterior roots of the spinal nerves. The anterior roots are formed by motor nerve fibers and transmit efferent impulses to the working organs. Sensory nerves predominate over motor nerves, so there is a primary analysis of the incoming afferent signals and the formation of reactions that are most important for the body in this moment(the transmission of numerous afferent impulses to a limited number of efferent neurons is called convergence).
Total amount spinal cord neurons is about 13 million. They are subdivided: 1) by department nervous system– neurons of somatic and autonomic NS; 2) by appointment - efferent, afferent, insertion; 3) by influence - excitatory and inhibitory.
Functions of neurons in the spinal cord.
Efferent neurons belong to the somatic nervous system and innervate skeletal muscles - motor neurons. There are alpha and gamma motor neurons. A-motor neurons carry out transmission of signals from the spinal cord to skeletal muscles. The axons of each motor neuron divide many times, so each of them covers many muscle fibers, forming a motor motor unit with it. G-motor neurons innervate the muscle fibers of the muscle spindle. They have a high frequency of impulses, receive information about the state of the muscle spindle through intermediate neurons (intercalary). Generate pulses with a frequency of up to 1000 per second. These are phonoactive neurons with up to 500 synapses on their dendrites.
Afferent neurons somatic NS are localized in the spinal ganglia and ganglia of the cranial nerves. Their processes conduct impulses from muscle, tendon, and skin receptors, enter the corresponding segments of the spinal cord, and connect by synapses with intercalary or alpha motor neurons.
Function intercalary neurons consists in the organization of communication between the structures of the spinal cord.
Neurons of the autonomic nervous system are intercalary . Sympathetic neurons located in the lateral horns of the thoracic spinal cord, they have a rare impulse frequency. Some of them are involved in maintaining vascular tone, others in the regulation of the smooth muscles of the digestive system.
The collection of neurons forms the nerve centers.
The spinal cord contains control centers most internal organs and skeletal muscles. Centers skeletal muscle control are located in all parts of the spinal cord and innervate, according to the segmental principle, the skeletal muscles of the neck (Ci-Civ), diaphragm (Ciii-Cv), upper limbs (Cv-Thii), trunk (Thiii-Li), lower limbs (Lii-Sv). When certain segments of the spinal cord or its pathways are damaged, specific motor and sensory disorders develop.
Functions of the spinal cord:
A) provides a two-way connection between the spinal nerves and the brain - a conductive function;
B) carries out complex motor and vegetative reflexes - a reflex function.
The spinal cord is the most ancient formation of the central nervous system; it first appears in the lancelet
A characteristic feature of the organization of the spinal cord is the periodicity of its structure in the form of segments with inputs in the form of posterior roots, a cell mass of neurons (gray matter) and outputs in the form of anterior roots.
The human spinal cord has 31-33 segments: 8 cervical, 12 thoracic, 5 lumbar, 5 sacral, 1-3 coccygeal.
Morphological boundaries between segments of the spinal cord do not exist. Each segment innervates three metameres of the body through its roots and also receives information from three metameres of the body. As a result, each metamere of the body is innervated by three segments and transmits signals to three segments of the spinal cord.
The posterior roots are afferent, sensory, centripetal, and the anterior roots are efferent, motor, centrifugal (Bell-Magendie law).
Afferent inputs to the spinal cord are organized by the axons of the spinal ganglia, which lie outside the spinal cord, and by the axons of the sympathetic and parasympathetic divisions of the autonomic nervous system.
The first group of afferent inputs of the spinal cord is formed by sensory fibers coming from muscle receptors, tendon receptors, periosteum, and joint membranes. This group of receptors forms the beginning of the so-called proprioceptive sensitivity.
The second group of afferent inputs of the spinal cord starts from skin receptors: pain, temperature, tactile, pressure.
The third group of afferent inputs of the spinal cord is represented by fibers from visceral organs, this is the viscero-receptive system.
Efferent (motor) neurons are located in the anterior horns of the spinal cord, and their fibers innervate the entire skeletal muscles.
Features of the neural organization of the spinal cord
The neurons of the spinal cord form its gray matter in the form of symmetrically located two anterior and two posterior horns. the nuclei, elongated along the length of the spinal cord, and on the transverse section are located in the shape of the letter H. In the thoracic region, the spinal cord has, in addition to those mentioned, also lateral horns.
The posterior horns perform mainly sensory functions; signals are transmitted from them to the overlying centers, to the structures of the opposite side, or to the anterior horns of the spinal cord.
In the anterior horns are neurons that give their axons to the muscles. All descending pathways of the central nervous system that cause motor responses end at the neurons of the anterior horns. In this regard, Sherrington called them "the common final path".
In the lateral horns, starting from the 1st thoracic segment of the spinal cord and up to the first lumbar segments, there are neurons of the sympathetic, and in the sacral - of the parasympathetic division of the autonomic nervous system.
The human spinal cord contains about 13 million neurons, of which 3% are motor neurons, and 97% are intercalary. Functionally, spinal cord neurons can be divided into 4 main groups:
1) motor neurons, or motor, - cells of the anterior horns, the axons of which form the anterior roots;
2) interneurons - neurons that receive information from the spinal ganglia and are located in the posterior horns. These neurons respond to pain, temperature, tactile, vibrational, proprioceptive stimuli;
3) sympathetic, parasympathetic neurons are located mainly in the lateral horns. The axons of these neurons exit the spinal cord as part of the anterior roots;
4) associative cells - neurons of the spinal cord's own apparatus, establishing connections within and between segments.
In the middle zone of the gray matter (between the posterior and anterior horns) of the spinal cord there is an intermediate nucleus (Cajal nucleus) with cells whose axons go up or down by 1-2 segments and give collaterals to the neurons of the ipsi- and contralateral side, forming a network. There is a similar network at the top of the posterior horn of the spinal cord - this network forms the so-called gelatinous substance (Roland's gelatinous substance) and performs the functions of the reticular formation of the spinal cord. The middle part of the gray matter of the spinal cord contains mainly short-axon spindle-shaped cells; , between the cells of its anterior and posterior horns.
Motoneurons. The axon of a motor neuron innervates hundreds of muscle fibers with its terminals, forming a motor neuron unit. Several motor neurons can innervate one muscle, in which case they form the so-called motor neuron pool. The excitability of motor neurons is different, therefore, with different intensity of stimulation, a different number of fibers of one muscle is involved in contraction. With the optimal strength of irritation, all fibers of this muscle are reduced; in this case, the maximum contraction develops. Motor neurons can generate impulses with a frequency of up to 200 per second.
Interneurons. These intermediate neurons, generating impulses with a frequency of up to 1000 per second, are background-active and have up to 500 synapses on their dendrites. The function of interneurons is to organize connections between the structures of the spinal cord and ensure the influence of ascending and descending pathways on the cells of individual segments of the spinal cord. A very important function of interneurons is the inhibition of neuron activity, which ensures the preservation of the direction of the excitation pathway. Excitation of interneurons associated with motor cells has an inhibitory effect on antagonist muscles.
The neurons of the sympathetic division of the autonomic nervous system are located in the lateral horns of the thoracic spinal cord, have a rare impulse frequency (3-5 per second), parasympathetic neurons are localized in the sacral spinal cord.
With irritation or lesions of the posterior roots, girdle pains are observed at the level of the metamer of the affected segment, sensitivity decreases, reflexes disappear or weaken. If an isolated lesion of the posterior horn occurs, pain and temperature sensitivity on the side of the injury is lost, while tactile and proprioceptive sensations are preserved, since axons of temperature and pain sensitivity go from the posterior root to the posterior horn, and axons of tactile and proprioceptive - directly to the posterior column and along the conductive paths rise up.
The defeat of the anterior horn and the anterior root of the spinal cord leads to paralysis of the muscles, which lose their tone, atrophy, and the reflexes associated with the affected segment disappear.
The defeat of the lateral horns of the spinal cord is accompanied by the disappearance of skin vascular reflexes, impaired sweating, trophic changes in the skin and nails. Bilateral damage to the parasympathetic department at the level of the sacrum leads to impaired defecation and urination.
1.1. Nervous system: general structure
The nervous system is a body system that combines and regulates various physiological processes in accordance with changing conditions of the external and internal environment. The nervous system consists of sensory components that respond to stimuli emanating from the environment, integrative components that process and store sensory and other data, and motor components that control movements and secretory activity of the glands.
The nervous system perceives sensory stimuli, processes information and forms behavior. Special types of information processing are learning and memory, due to which, when the environment changes, behavior adapts taking into account previous experience. Other systems such as the endocrine and immune systems are also involved in these functions, but the nervous system is specialized to carry out specified functions. Information processing refers to the transmission of information in neural networks, the transformation of signals by combining them with other signals (neural integration), the storage of information in memory and the retrieval of information from memory, the use of sensory information for perception, thinking, learning, planning (preparation) and the execution of motor commands, the formation of emotions. Interactions between neurons are carried out through both electrical and chemical processes.
Behavior is a complex of reactions of the organism to changing conditions of the external and internal environment. Behavior can be a purely internal, hidden process (cognition) or accessible to external observation (motor or vegetative reactions). In humans, a set of those behavioral acts that are associated with speech is especially important. Each reaction, simple or complex, is provided by nerve cells organized into neural networks (nerve ensembles and pathways).
The nervous system is divided into central and peripheral (Fig. 1.1). The central nervous system (CNS) consists of the brain and spinal cord. The peripheral nervous system includes roots, plexuses and nerves.
Rice. 1.1. General structure of the nervous system.
BUT- Central nervous system. B- Brain stem: 1 - telencephalon; 2 - diencephalon; 3 - midbrain; 4 - bridge and cerebellum, 5 - medulla oblongata, 6 - telencephalon median structures. IN- Spinal cord: 7 - spinal cone; 8 - terminal threads. G- Peripheral nervous system: 9 - ventral root; 10 - dorsal spine; 11 - spinal ganglion; 12 - spinal nerve; 13 - mixed peripheral nerve; 14 - epineurium; 15 - perineurium; 16 - myelin nerve; 17 - fibrocyte; 18 - endoneurium; 19 - capillary; 20 - unmyelinated nerve; 21 - skin receptors; 22 - the end of the motor neuron; 23 - capillary; 24 - muscle fibers; 25 - the nucleus of the Schwann cell; 26 - interception of Ranvier; 27 - sympathetic trunk; 28 - connecting branch
central nervous system
The central nervous system collects and processes information about the environment coming from the receptors, forms reflexes and other behavioral reactions, plans and carries out arbitrary movements. In addition, the central nervous system provides the so-called higher cognitive (cognitive) functions. In the central nervous system, processes associated with memory, learning and thinking take place.
The brain in the process of ontogenesis is formed from cerebral vesicles resulting from uneven growth of the anterior sections of the medullary tube (Fig. 1.2). These bubbles form the forebrain (prosencephalon) midbrain (mesencephalon) and rhomboid brain (rhombencephalon). In the future, the end brain is formed from the forebrain (telencephalon) and intermediate (diencephalon) brain, and the rhomboid brain is divided into the posterior (metencephalon) and oblong (myelencephalon, or medulla oblongata) brain. From the telencephalon, respectively, the cerebral hemispheres, basal ganglia are formed, from the diencephalon - the thalamus, epithalamus, hypothalamus, metathalamus, optic tracts and nerves, retina. The optic nerves and retina are parts of the central nervous system, as if taken out of the brain. From the midbrain, the plate of the quadrigemina and the legs of the brain are formed. The pons and cerebellum form from the hindbrain. The pons of the brain borders below on the medulla oblongata.
The back of the medullary tube forms the spinal cord, and its cavity becomes the central canal of the spinal cord. The spinal cord consists of cervical, thoracic, lumbar, sacral and coccygeal regions, each of which in turn consists of segments.
The CNS is divided into gray and white matter. The gray matter is a cluster of neuron bodies, the white matter is the processes of neurons covered with a myelin sheath. In the brain, gray matter is located in the cerebral cortex, in the subcortical ganglia, the nuclei of the brain stem, the cerebellar cortex and its nuclei. In the spinal cord, gray matter is concentrated in its middle, white - on the periphery.
Peripheral nervous system
The peripheral nervous system (PNS) is responsible for the interface between the environment (or excitable cells) and the CNS. The PNS includes sensory (receptors and primary afferent neurons) and motor (somatic and autonomic motor neurons) components.
 Rice. 1.2. Embryonic development of the mammalian nervous system. Scheme of the development of the neural felling at stage three (BUT) and five (B) brain bubbles. A. I- General form side: 1 - cranial bend; 2 - cervical bend; 3 - spinal node. II- Top view: 4 - forebrain; 5 - midbrain; 6 - diamond-shaped brain; 7 - neurocoel; 8 - wall of the neural tube; 9 - rudimentary spinal cord.
Rice. 1.2. Embryonic development of the mammalian nervous system. Scheme of the development of the neural felling at stage three (BUT) and five (B) brain bubbles. A. I- General form side: 1 - cranial bend; 2 - cervical bend; 3 - spinal node. II- Top view: 4 - forebrain; 5 - midbrain; 6 - diamond-shaped brain; 7 - neurocoel; 8 - wall of the neural tube; 9 - rudimentary spinal cord.
B. I- General side view. B. II- Top view: 10 - telencephalon; 11 - lateral ventricle; 12 - diencephalon; 13 - eye stalk; 14 - lens; 15 - optic nerve; 16 - midbrain; 17 - hindbrain; 18 - medulla oblongata; 19 - spinal cord; 20 - central channel; 21 - fourth ventricle; 22 - aqueduct of the brain; 23 - third ventricle. III- Side view: 24 - new bark; 25 - interventricular septum; 26 - striatum; 27 - pale ball; 28 - hippocampus; 29 - thalamus; 30 - pineal body; 31 - upper and lower mounds; 32 - cerebellum; 33 - hindbrain; 34 - spinal cord; 35 - medulla oblongata; 36 - bridge; 37 - midbrain; 38 - neurohypophysis; 39 - hypothalamus; 40 - amygdala; 41 - olfactory tract; 42 - olfactory cortex
Sensory part of the PNS. Sensory perception is the conversion of the energy of an external stimulus into a nerve signal. It is carried out by specialized structures - receptors that perceive the impact on the body of various types of external energy, including mechanical, light, sound, chemical stimuli, temperature changes. The receptors are located at the peripheral endings of the primary afferent neurons, which transmit the received information to the central nervous system along the sensory fibers of the nerves, plexuses, spinal nerves, and, finally, along the posterior roots of the spinal cord (or along the cranial nerves). The cell bodies of the dorsal roots and cranial nerves are located in the spinal (spinal) ganglia or in the ganglia of the cranial nerves.
The motor part of the PNS. The motor component of the PNS includes somatic and autonomic (autonomous) motor neurons. Somatic motor neurons innervate striated muscles. The cell bodies are located in the anterior horns of the spinal cord or in the brainstem, they have long dendrites that receive many synaptic inputs. The motor neurons of each muscle make up a specific motor nucleus - a group of CNS neurons that have similar functions. For example, facial muscles are innervated from the nucleus of the facial nerve. Axons of somatic motor neurons leave the CNS through the anterior root or through the cranial nerve.
Autonomic (autonomous) motor neurons send nerves to smooth muscle fibers and glands - preganglionic and postganglionic neurons of the sympathetic and parasympathetic nervous system. Preganglionic neurons are located in the central nervous system - in the spinal cord or in the brain stem. Unlike somatic motor neurons, autonomic preganglionic neurons form synapses not on effector cells (smooth muscle or glands), but on postganglionic neurons, which in turn synaptically contact directly with effectors.
1.2. Microscopic structure of the nervous system
The basis of the nervous system are nerve cells, or neurons, specialized in receiving incoming signals and in transmitting signals to other neurons or effector cells. In addition to nerve cells, the nervous system contains glial cells and connective tissue elements. Cells of neuroglia (from the Greek "glia" - glue) are
perform supporting, trophic, regulatory functions in the nervous system, participating in almost all types of neuronal activity. Quantitatively, they prevail over neurons and occupy the entire volume between the vessels and nerve cells.
Nerve cell
The main structural and functional unit of the nervous system is the neuron (Fig. 1.3). In a neuron, a body (soma) and processes are distinguished: dendrites and an axon. The soma and dendrites represent the receptive surface of the cell. The axon of a nerve cell forms synaptic connections with other neurons or with effector cells. A nerve impulse always propagates in one direction: along the dendrites to the cell body, along the axon - from the cell body (Ramon y Cajal's law of dynamic polarization of the nerve cell). As a rule, a neuron has many "inputs" carried out by dendrites, and only one "output" (axon) (see Fig. 1.3).
Neurons communicate with each other using action potentials that propagate along the axons. Action potentials are transmitted from one neuron to the next as a result of synaptic transmission. An action potential reaching the presynaptic end usually triggers the release of a neurotransmitter, which either excites the postsynaptic cell so that a discharge of one or more action potentials occurs in it, or inhibits its activity. Axons not only transmit information in nerve
 Rice. 1.3. The structure of a neuron. BUT- A typical neuron, consisting of the body itself, dendrites and an axon: 1 - the beginning of the axon; 2 - dendrites; 3 - neuron body; 4 - axon; 5 - Schwann cage; 6 - branching of the axon. B- Enlarged neuron body. The axonal hillock does not contain Nissl's substance: 7 - core; 8 - Golgi apparatus; 9 - mitochondria; 10 - axonal hillock; 11 - Nissl substance
Rice. 1.3. The structure of a neuron. BUT- A typical neuron, consisting of the body itself, dendrites and an axon: 1 - the beginning of the axon; 2 - dendrites; 3 - neuron body; 4 - axon; 5 - Schwann cage; 6 - branching of the axon. B- Enlarged neuron body. The axonal hillock does not contain Nissl's substance: 7 - core; 8 - Golgi apparatus; 9 - mitochondria; 10 - axonal hillock; 11 - Nissl substance
chains, but also deliver chemicals by axon transport to synaptic endings.
There are numerous classifications of neurons according to the shape of their body, the length and shape of the dendrites, and other features (Fig. 1.4). According to their functional significance, nerve cells are divided into afferent (sensory, sensory), delivering impulses to the center, efferent (motor, motor), carrying information from the center to the periphery, and interneurons, in which impulses are processed and collateral connections are organized.
A nerve cell performs two main functions: specific processing of incoming information and transmission of a nerve impulse, and biosynthetic, aimed at maintaining its vital activity. This finds expression in the ultrastructure of the nerve cell. The transfer of information from one nerve cell to another, the unification of nerve cells into systems and complexes of varying complexity are carried out by neuron structures: axons, dendrites and synapses. Organelles associated with the provision of energy metabolism, the protein-synthesizing function of the cell, are found in most cells; in nerve cells, they perform the functions of energy supply to the cell, processing and transmission of information (see Fig. 1.3).
Structure of a neuron. Soma. The body of the nerve cell has a round or oval shape, in the center (or slightly eccentric) is the nucleus. It contains the nucleolus and is surrounded by outer and inner nuclear membranes about 70 Å thick each, separated by peri-
 Rice. 1.4. Variants of neurons of different shapes.
Rice. 1.4. Variants of neurons of different shapes.
BUT- Pseudo-unipolar neuron. B- Purkinje cell (dendrites, axon). IN- pyramidal cell (axon). G- motoneuron of the anterior horn (axon)
nuclear space, the dimensions of which are variable. In the karyoplasm, lumps of chromatin are distributed, localized mainly at the inner nuclear membrane. In the cytoplasm of nerve cells there are elements of a granular and non-granular cytoplasmic reticulum, polysomes, ribosomes, mitochondria, lysosomes, multibubble bodies and other organelles (Fig. 1.5).
The apparatus of biosynthesis in neurons includes Nissl bodies - flattened cisterns of the granular endoplasmic reticulum tightly adjacent to each other, as well as a well-defined Golgi apparatus. In addition, the soma contains numerous mitochondria, which determine its energy metabolism, and elements of the cytoskeleton, including neurofilaments and microtubules. Lysosomes and phagosomes are the main organelles of the "intracellular digestive tract".
Dendrites. Dendrites and their branches determine the receptive field of a particular cell (see Fig. 1.5). An electron microscopic examination reveals that the body of a neuron gradually turns into a dendrite. There is no sharp boundary and pronounced differences in the ultrastructure of the soma and the initial section of a large dendrite. Dendrites are very variable in shape, size, branching, and ultrastructure. Usually several dendrites extend from the cell body. The length of the dendrite can exceed 1 mm, they account for more than 90% of the surface area of the neuron.
The main components of the dendritic cytoplasm are microtubules and neurofilaments; the proximal parts of the dendrites (closer to the cell body) contain Nissl bodies and sections of the Golgi apparatus. Previously, it was believed that dendrites are electrically non-excitable; it has now been proven that the dendrites of many
 Rice. 1.5. Ultrastructure of the nerve cell.
Rice. 1.5. Ultrastructure of the nerve cell.
1 - core; 2 - granular endoplasmic reticulum; 3 - lamellar complex (Golgi); 4 - mitochondria; 5 - lysosomes; 6 - multivesicular body; 7 - polysomes
neurons have voltage-dependent conductivity, which is due to the presence of calcium channels on their membranes, upon activation of which action potentials are generated.
Axon. The axon originates at the axon hillock - a specialized section of the cell (usually the soma, but sometimes the dendrite) (see Fig. 1.3). The axon and axon hillock differ from the soma and proximal portions of dendrites by the absence of a granular endoplasmic reticulum, free ribosomes, and the Golgi apparatus. The axon contains a smooth endoplasmic reticulum and a pronounced cytoskeleton.
Axons are covered with a myelin sheath, forming myelin fibers. Bundles of fibers (which may contain individual unmyelinated fibers) make up the white matter of the brain, cranial and peripheral nerves. When the axon passes into the presynaptic ending filled with synaptic vesicles, the axon forms a cone-shaped extension.
Interweaving of axons, dendrites and processes of glial cells create complex, non-repeating patterns of the neuropil. The distribution of axons and dendrites, their relative positions, afferent-efferent relationships, patterns of synaptoarchitectonics determine the mechanisms of the integrative function of the brain.
Types of neurons. Polymorphism in the structure of neurons is determined by their different role in the systemic activity of the brain as a whole. Thus, the neurons of the ganglia of the posterior roots of the spinal cord (spinal ganglia) receive information not through synaptic transmission, but from sensory nerve endings in the receptor organs. In accordance with this, the cell bodies of these neurons are devoid of dendrites and do not receive synaptic endings (bipolar cells; Fig. 1.6). After leaving the cell body, the axon of such a neuron is divided into two branches, one of which (the peripheral process) is sent as part of the peripheral nerve to the receptor, and the other branch (the central process) enters the spinal cord (as part of the posterior root) or the brainstem ( within the cranial nerve). Other types of neurons, such as pyramidal cells in the cerebral cortex and Purkinje cells in the cerebellar cortex, are busy processing information. Their dendrites are covered with dendritic spines and have an extensive surface; they receive a huge number of synaptic inputs (multipolar cells; see Fig. 1.4, 1.6). One can classify neurons according to the length of their axons. In Golgi type 1 neurons, axons are short, ending, like dendrites, close to the soma. Neurons of the 2nd type have long axons, sometimes longer than 1 m.
neuroglia
Another group of cellular elements of the nervous system is neuroglia (Fig. 1.7). In the human CNS, the number of neuroglial cells is an order of magnitude greater than the number of neurons: 10 13 and 10 12, respectively. A close morphological relationship is the basis for physiological and pathological interactions between glia and neurons. Their relationship is described by the concept of dynamic neuronal-glial signaling processes. The ability to transmit signals from neurons to glia, and thus to other neurons, opens up many options for intercellular "cross-talk".
There are several types of neuroglia; in the CNS, neuroglia are represented by astrocytes and oligodendrocytes, and in the PNS by Schwann cells and satellite cells. In addition, microglial cells and ependymal cells are considered central glial cells.
Astrocytes(named due to their stellate shape) regulate the state of the microenvironment around CNS neurons. Their processes surround groups of synaptic endings, which are isolated from neighboring synapses as a result. Special processes - "legs" of astrocytes form contacts with capillaries and connective tissue on the surface of the brain and spinal cord (pia mater) (Fig. 1.8). Legs limit the free diffusion of substances in the CNS. Astrocytes can actively absorb K+ and neurotransmitters and then metabolize them. Due to selectively increased permeability for K+ ions, astroglia regulates the activation of enzymes necessary to maintain the metabolism of neurons, as well as to remove mediators and other agents released during neuro-
 Rice. 1.6. Classification of neurons according to the number of processes extending from the cell body.
Rice. 1.6. Classification of neurons according to the number of processes extending from the cell body.
BUT - bipolar. B- pseudo-unipolar. IN- multipolar. 1 - dendrites; 2 - axon
 Rice. 1.7. Main types of glial cells.
Rice. 1.7. Main types of glial cells.
BUT- Protoplasmic astrocyte. B- microglial cell. IN- oligoderdrocyte. G- fibrous astrocyte
nal activity. Astroglia is involved in the synthesis of immune mediators: cytokines, other signaling molecules (cyclic guanosine monophosphate - coMP, nitric oxide - NO), then transferred to neurons - in the synthesis of glial growth factors ( GDNF), involved in trophism and repair of neurons. Astrocytes are able to respond to an increase in the synaptic concentration of neurotransmitters and a change in the electrical activity of neurons by changes in the intracellular concentration of Ca 2+ . This creates a "wave" of Ca 2+ migration between astrocytes, capable of modulating the state of many neurons.
Thus, astroglia, not being only a trophic component of the nervous system, is involved in the specific functioning of the nervous tissue. In the cytoplasm of astrocytes there are glial filaments that perform a mechanical support function in the CNS tissue. In case of damage, the processes of astrocytes containing glial filaments undergo hypertrophy and form a glial scar.
main function oligodendrocytes is to provide electrical insulation of axons by forming a myelin sheath (Fig. 1.9). It is a multi-layered wrapping spirally wound over the plasma membrane of axons. In the PNS, the myelin sheath is formed by the membranes of Schwann cells (see Figure 1.18). Myelin Presents
It is a package of sheets of specific plasma membranes rich in phospholipids, and also contains several types of proteins, different in the CNS and PNS. Protein structures allow plasma membranes to pack tightly together. With the growth of the glial cell membrane, it rotates around the axon of the neuron with the formation of a layered spiral with a double plasma membrane around the axon. The thickness of the myelin sheath can be 50-100 membranes, which play the role of an electrical insulator of the axon, preventing ion exchange between the axon cytosol and the extracellular environment.
In addition, the neuroglia include satellite cells that encapsulate the neurons of the ganglia of the spinal and cranial nerves, regulating the microenvironment around these neurons in the same way that astrocytes do (Fig. 1.10).
Another type of cell microglia, or latent phagocytes. Microglia is the only representation of immunocompetent cells in the CNS. It is widely represented in all human brain tissue and makes up 9-12% of the total glial population in the gray matter and 7.5-9% in the white matter. Unlike astrocytes, microglial cells originate from stem cells and, under normal conditions, have branched
 Rice. 1.8. Interaction of astrocytes with surrounding cellular elements.
Rice. 1.8. Interaction of astrocytes with surrounding cellular elements.
1 - tanycite; 2 - cavity of the ventricle; 3 - ependymal cells; 4 - capillary; 5 - neuron; 6 - myelinated axon; 7 - pia mater; 8 - subarachnoid space.
The figure shows two astrocytes and their relationship with the ependymal cells lining the ventricle, perikaryon, neuron dendrites, capillary, and pia mater squamous epithelium. It should be noted that this figure is schematic and the connection of the neuron simultaneously with the ventricle and subarachnoid space is unlikely.
 Rice. 1.9. Oligodendrocyte: formation of the myelin sheath of the axon. 1 - axon; 2 - myelin; 3 - smooth endoplasmic reticulum; 4 - neurofilaments; 5 - mitochondria
Rice. 1.9. Oligodendrocyte: formation of the myelin sheath of the axon. 1 - axon; 2 - myelin; 3 - smooth endoplasmic reticulum; 4 - neurofilaments; 5 - mitochondria
 Rice. 1.10. Interaction of glial cells and neurons. Schematically shown by arrows. 1 - satellite glial cell; 2 - glial cell synthesizing myelin
Rice. 1.10. Interaction of glial cells and neurons. Schematically shown by arrows. 1 - satellite glial cell; 2 - glial cell synthesizing myelin
viscous form with many processes. Activation of microglia, in particular under conditions of hypoxia, is accompanied by the production of pro-inflammatory mediators with toxic properties. The chronic inflammatory reaction they maintain in the brain tissue leads to delayed neuronal losses, microcirculatory disorders, and changes in the function of the blood-brain barrier.
Under pathological conditions, microglial cells retract their processes and take on an amoeboid shape, which corresponds to their pronounced functional activation up to the state of phagocytosis. When the brain tissue is damaged, microglia, along with phagocytes penetrating the CNS from the bloodstream, contributes to the removal of cellular decay products.
The CNS tissue is separated from the cerebrospinal fluid (CSF) that fills the ventricles of the brain by an epithelium that is formed by ependymal cells. The ependyma mediates the diffusion of many substances between the extracellular space of the brain and the CSF. CSF is secreted by specialized ependymal cells of the choroid plexuses in the ventricular system.
The supply of nutrients to the brain cells and the removal of cell waste products occur through the vascular
system. Although nervous tissue is replete with capillaries and other blood vessels, the blood-brain barrier (BBB) limits the diffusion of many substances between the blood and CNS tissue.
1.3. Electrical transmission of information between neurons
The normal activity of the nervous system depends on the excitability of its neurons. Excitability- this is the ability of cell membranes to respond to the action of adequate stimuli with specific changes in ionic conductivity and membrane potential. Excitation- an electrochemical process that occurs exclusively on the cytoplasmic membrane of the cell and is characterized by changes in its electrical state, which triggers a function specific to each tissue. Thus, the excitation of the muscle membrane causes its contraction, and the excitation of the neuron membrane causes the conduction of an electrical signal along the axons. Neurons have not only voltage-controlled, i.e. ion channels regulated by the action of an electrical exciter, but also chemocontrolled and mechanically controlled.
There are differences in the relationship between membrane potential/membrane permeability and the type of stimulus. When exposed to an electrical stimulus, the chain of events is as follows: stimulus (electric current) => shift of the membrane potential (to the critical potential) => activation of voltage-gated ion channels => change in the ionic permeability of the membrane => change in ion currents through the membrane => further shift in the membrane potential (formation of an action potential).
When exposed to a chemical stimulus, a fundamentally different chain of events occurs: stimulus (chemical substance) => chemical binding of the stimulus and the chemo-controlled ion channel receptor => change in the conformation of the ligand-receptor complex and opening of receptor-controlled (chemo-controlled) ion channels => change in the ionic permeability of the membrane => change in ion currents through the membrane => shift in the membrane potential (formation, e.g. local potential).
The chain of events under the influence of a mechanical stimulus is similar to the previous one, since in this case the receptors are also activated.
gated ion channels: stimulus (mechanical stress) => change in membrane tension => opening of receptor-controlled (mechanically controlled) ion channels => change in membrane ion permeability => change in ion currents through the membrane => shift in membrane potential (formation of mechanically induced potential).
The passive electrical properties of a cell are related to the electrical properties of its membrane, cytoplasm, and external environment. Electrical Properties The cell membrane is determined by its capacitive and resistive characteristics, since the lipid bilayer can be directly likened to both a capacitor and a resistor. The capacitive characteristics of the lipid bilayer and the real membrane are similar, while the resistive ones differ due to the presence, first of all, of proteins that form ion channels. For most cells, the input resistance behaves non-linearly: for current flowing in one direction, it is greater than for the opposite direction. This property of asymmetry reflects an active reaction and is called straightening. The current flowing through the membrane is determined by the capacitive and resistive components. The resistive component describes the actual ionic current, since electricity is carried in the cell by ions. The movement of ions into or out of the cell is prevented by the plasma membrane. Because the membrane is a lipid bilayer that is impermeable to ions, it is resistant. Conversely, the membrane has some conductivity for the ions that pass through the ion channels. Due to the obstacle to the free movement of ions, the same ions are found outside and inside the cell, but in different concentrations.
There are two fundamental mechanisms for the movement of substances through the membrane - through simple diffusion (Fig. 1.11) and when
 Rice. 1.11. Transport of substances across the cell membrane.
Rice. 1.11. Transport of substances across the cell membrane.
BUT- simple diffusion. B- facilitated diffusion. IN- active transport: 1- membrane
the power of specific carriers built into the membrane and representing transmembrane integral proteins. The latter mechanism includes facilitated diffusion and active ion transport, which can be primary active and secondary active.
Through simple diffusion (without the help of a carrier), water-insoluble organic compounds and gases (oxygen and carbon dioxide) can be transported through the lipid bilayer by dissolving them in the lipids of the cell membrane; ions Na +, Ca 2+, K +, Cl - through the ion channels of the cell membrane, connecting the cytoplasm of cells with the external environment (passive ion transport, which is determined by an electrochemical gradient and is directed from a larger electrochemical potential to a smaller one: inside the cell for Na + ions, Ca 2+, Cl -, outside - for K + ions); water molecules through the membrane (osmosis).
With the help of specific carriers, energy-independent facilitated diffusion of a number of compounds is carried out (see Fig. 1.11). A striking example of facilitated diffusion is the transport of glucose across the neuron membrane. Without a specialized astrocytic transporter, the entry of glucose into neurons would be practically impossible, since it is a relatively large polar molecule. Due to its rapid conversion to glucose-6-phosphate, the intracellular glucose level is lower than the extracellular level, and thus a gradient is maintained to ensure a continuous flow of glucose into neurons.
The energy-dependent primary active transport of Na+, Ca2+, K+, and H+ ions is the energy-dependent transport of substances against their electrochemical gradients (see Fig. 1.11). Thanks to him, cells can accumulate ions in concentrations higher than in the environment. The movement from a lower to a higher concentration and the maintenance of a steady-state gradient are possible only with continuous energy supply to the transport process. Primary active transport involves direct consumption of ATP. ATP energy pumps (ATPase) transport ions against their concentration gradient. Based on the characteristics of the molecular organization, 3 classes are distinguished - P, V and F (Fig. 1.12). All three classes of ATPases have one or more ATP binding sites on the cytosolic membrane surface. Class P includes Ca 2+ -ATPase and Na + /K + -ATPase. Active ion transport carriers are specific for the transported substance and are saturable, i.e. their flux is maximum when all specific sites of binding to the carried substance are occupied.
Many gradients of the electrochemical potential of the cell, which are a necessary condition for the passive transport of ions, appear as a result of their active transport. Thus, K + and Na + gradients arise as a result of their active transfer of Na + / K + - by the pump (Fig. 1.13). Due to the activity of Na + /K + -pump inside the cell, K + ions are present in a higher concentration, but they tend to pass through diffusion into the extracellular medium along the concentration gradient. To maintain the equality of positive and negative charges inside the cell, the release of K + ions into the external environment must be compensated by the entry of Na + ions into the cell. Since the membrane at rest is much less permeable to Na + ions than to K + ions, potassium must leave the cell along a concentration gradient. As a result, a positive charge accumulates on the outside of the membrane, and a negative charge accumulates on the inside. This maintains the resting potential of the membrane.
The secondary active transport of a number of ions and molecules also uses the energy accumulated as a result of ATP consumption and spent on creating a concentration gradient. The ion concentration gradient relative to the membrane is used as an energy source created by primary active transport (Fig. 1.14). Thus, secondary active transport includes cotransport and countertransport: the flow of ions from a higher (higher energy state) to a lower (lower energy state) concentration provides the energy to move the actively transported substance from the area of its low concentration to the area of high.
 Rice. 1.12. Three classes of ATP dependent ion pumps. BUT- P-class. B- F 1 - class IN- V 1 -class
Rice. 1.12. Three classes of ATP dependent ion pumps. BUT- P-class. B- F 1 - class IN- V 1 -class
Cell potentials determined by passive ion transport
In response to subthreshold, close to the threshold and threshold electric current impulses, a passive electrotonic potential, a local response and an action potential arise, respectively (Fig. 1.15). All these potentials are determined by passive ion transport across the membrane. Their occurrence requires polarization of the cell membrane, which can be carried out extracellularly (usually observed on nerve fibers) and intracellularly (usually noted on the cell body).
Passive electrotonic potential arises in response to a subthreshold impulse, which does not lead to the opening of ion channels and is determined only by the capacitive and resistive properties of the cell membrane. The passive electrotonic potential is characterized by a time constant, which reflects the passive properties of the membrane, the time course of changes in the membrane potential, i.e. the rate at which it changes from one value to another. Pass-
 Rice. 1.13. Working mechanism of Na + /K + pump
Rice. 1.13. Working mechanism of Na + /K + pump
 Rice. 1.14. The mechanism of operation of secondary-active transport. BUT- Stage 1. B- Stage 2. IN- Step 3: 1 - Na+; 2 - the molecule of the substance to be transferred against the concentration gradient; 3 - conveyor. When Na + binds to the carrier, allosteric changes occur in the binding site of the carrier protein for the molecule of the transferred substance, which causes a conformational change in the carrier protein, allowing Na + ions and the bound substance to exit on the other side of the membrane
Rice. 1.14. The mechanism of operation of secondary-active transport. BUT- Stage 1. B- Stage 2. IN- Step 3: 1 - Na+; 2 - the molecule of the substance to be transferred against the concentration gradient; 3 - conveyor. When Na + binds to the carrier, allosteric changes occur in the binding site of the carrier protein for the molecule of the transferred substance, which causes a conformational change in the carrier protein, allowing Na + ions and the bound substance to exit on the other side of the membrane
A strong electrotonic potential is characterized by the equality of the rates of rise and fall of the exponent. There is a linear relationship between the amplitudes of the electrical stimulus and the passive electrotonic potential, and an increase in the pulse duration does not change this pattern. Passive electrotonic potential propagates along the axon with attenuation, which is determined by the constant length of the membrane.
When the strength of the electric impulse approaches the threshold value, local membrane response, which is manifested by a change in the shape of the passive electrotonic potential and the development of an independent peak of small amplitude, resembling an S-shaped curve in shape (see Fig. 1.15). The first signs of a local response are recorded under the action of stimuli that are approximately 75% of the threshold value. With an increase in the irritating current, the amplitude of the local response increases non-linearly and can not only reach the critical potential, but also exceed it, without developing, however, into an action potential. The independent development of a local response is associated with an increase in the sodium permeability of the membrane through sodium channels that provide an incoming current, which, at a threshold stimulus, causes the depolarization phase of the action potential. However, with a subthreshold stimulus, this increase in permeability is insufficient to trigger the process of regenerative membrane depolarization, since only a small part of the sodium channels open. Started de-
 Rice. 1.15. cell membrane potentials.
Rice. 1.15. cell membrane potentials.
BUT- Dynamics of changes in the membrane potential depending on the strength of the depolarizing electric current impulse. B- Discrete increase in the strength of the depolarizing impulse
polarization stops. As a result of the release of K + ions from the cell, the potential returns to the level of the resting potential. Unlike the action potential, the local response does not have a clear threshold of occurrence and does not obey the all-or-nothing law: with an increase in the strength of the electrical impulse, the amplitude of the local response increases. In the body, the local response is the electrophysiological expression of local excitation and usually precedes the action potential. Sometimes a local response can exist on its own in the form of an excitatory postsynaptic potential. Examples of the independent value of the local potential are the conduction of excitation from amacrine cells of the retina - CNS neurons lacking axons, to synaptic endings, as well as the response of the postsynaptic membrane of a chemical synapse and the communicative transmission of information between nerve cells that generate synaptic potentials.
At the threshold value of the irritating electrical impulse, action potential, consisting of phases of depolarization and repolarization (Fig. 1.16). The action potential begins as a result of the displacement of the resting potential (for example, from -90 mV) to the level of the critical potential (different for different types of cells) under the action of a rectangular electric current pulse. The depolarization phase is based on the activation of all voltage-gated sodium channels, followed by
 Rice. 1.16. Changes in the membrane potential of a neuron (BUT) and conductivity of ions through the plasmalemma (B) when an action potential occurs. 1 - fast depolarization; 2 - overshoot; 3 - repolarization; 4 - threshold potential; 5 - hyperpolarization; 6 - resting potential; 7 - slow depolarization; 8 - action potential; 9 - permeability for sodium ions; 10 - permeability for potassium ions.
Rice. 1.16. Changes in the membrane potential of a neuron (BUT) and conductivity of ions through the plasmalemma (B) when an action potential occurs. 1 - fast depolarization; 2 - overshoot; 3 - repolarization; 4 - threshold potential; 5 - hyperpolarization; 6 - resting potential; 7 - slow depolarization; 8 - action potential; 9 - permeability for sodium ions; 10 - permeability for potassium ions.
Ion conduction curves are interconnected with the action potential curve
As a result, the passive transport of Na + ions into the cell increases and a shift in the membrane potential occurs up to 35 mV (this peak level is different for cells of different types). The excess of the action potential above the zero line is called an overshoot. Upon reaching the peak, the potential value falls into the negative region, reaching the resting potential (repolarization phase). Repolarization is based on the inactivation of voltage-gated sodium channels and the activation of voltage-gated potassium channels. Ions K + pass out of the cell by passive transport and the resulting current leads to a shift in the membrane potential to the negative region. The repolarization phase ends with trace hyperpolarization or trace depolarization - alternative ionic mechanisms for returning the membrane potential to the level of the resting potential (see Fig. 1.16). With the first mechanism, repolarization reaches a resting value and continues further into a more negative area, after which it returns to the level of the resting potential (trace hyperpolarization); in the second, repolarization occurs slowly and smoothly passes into the resting potential (trace depolarization). The development of the action potential is accompanied by phase changes in cell excitability - from increased excitability to absolute and relative refractoriness.
Bioelectric activity of neurons
The first type of bioelectrical activity of cells is inherent in silent neurons, which are not capable of independently generating action potentials. The resting potential of these cells does not change (Fig. 1.17).
Neurons of the second type are able to independently generate action potentials. Among them, cells are distinguished that generate regular and irregular rhythmic or burst (a burst consists of several action potentials, after which a short period of rest is observed) activity.
The third type of bioelectrical activity includes neurons that can independently generate fluctuations of the resting potential of a sinusoidal or sawtooth shape that do not reach the critical potential. Only rare oscillations can reach the threshold and cause the generation of single action potentials. These neurons are called pacemaker neurons (Fig. 1.17).
The "behavior" of individual neurons and interneuronal interactions are influenced by long-term polarization (depolarization or hyperpolarization) of postsynaptic cell membranes.
Stimulation of neurons with a constant depolarizing electrical current causes responses with rhythmic discharges of action potentials. After the cessation of prolonged depolarization of the membrane, post-activation inhibition in which the cell is unable to generate action potentials. The duration of the stage of post-activation inhibition directly correlates with the amplitude of the stimulating current. Then the cell gradually restores the usual rhythm of potential generation.
On the contrary, a constant hyperpolarizing current inhibits the development of the action potential, which is of particular importance in relation to neurons with spontaneous activity. An increase in cell membrane hyperpolarization leads to a decrease in the frequency of spike activity and an increase in the amplitude of each action potential; the next stage is the complete cessation of potential generation. After the cessation of prolonged hyperpolarization of the membrane, the phase begins post-brake activation, when the cell begins to spontaneously generate at a higher frequency than usual, action potentials. The duration of the post-activation activation stage directly correlates with the amplitude of the hyperpolarizing current, after which the cell gradually restores the usual rhythm of potential generation.
 Rice. 1.17. Types of bioelectrical activity of nerve cells
Rice. 1.17. Types of bioelectrical activity of nerve cells
1.4. Conduction of excitation along the nerve fiber
The patterns of conduction of excitation along nerve fibers are determined by both electrical and morphological features of axons. Nerve trunks are composed of myelinated and unmyelinated fibers. The membrane of an unmyelinated nerve fiber is in direct contact with the external environment, i.e. the exchange of ions between the intracellular and extracellular environment can occur at any point of the unmyelinated fiber. The myelinated nerve fiber is covered for a greater extent by a fatty (myelin) sheath that acts as an insulator (see Fig. 1.18).
Myelin from one glial cell forms a region of myelinated nerve fiber, separated from the next region formed by another glial cell, an unmyelinated area - the Ranvier interception (Fig. 1.19). The length of the node of Ranvier is only 2 µm, and the length of the myelinated fiber section between adjacent nodes of Ranvier reaches 2000 µm. Nodes of Ranvier are completely free of myelin and can come into contact with extracellular fluid, i.e. the electrical activity of the myelinated nerve fiber is limited by the Ranvier interception membrane, through which ions are able to penetrate. In these regions of the membrane, the highest density of voltage-gated sodium channels is noted.
The passive electrotonic potential propagates along the nerve fiber over short distances (Fig. 1.20), while its amplitude
 Rice. 1.18. Scheme of myelination of a peripheral nerve fiber. BUT- Stages of myelination. a - the axon is clasped by the process of the Schwann cell; b - the process of the Schwann cell winds around the axon; c - Schwann cell loses most of the cytoplasm, turning into a lamellar sheath around the axon. B- Unmyelinated axons surrounded by a Schwann cell process
Rice. 1.18. Scheme of myelination of a peripheral nerve fiber. BUT- Stages of myelination. a - the axon is clasped by the process of the Schwann cell; b - the process of the Schwann cell winds around the axon; c - Schwann cell loses most of the cytoplasm, turning into a lamellar sheath around the axon. B- Unmyelinated axons surrounded by a Schwann cell process
 Rice. 1.19. Structure of Ranvier interception.
Rice. 1.19. Structure of Ranvier interception.
1 - axon plasma membrane;
2 - myelin membranes; 3 - cytosol of the Schwann cell; 4 - Ranvier interception zone; 5 - plasma membrane of the Schwann cell
there, the rate of rise and fall decreases with distance (excitation decay phenomenon). The propagation of excitation in the form of an action potential is not accompanied by a change in the shape or amplitude of the potential, since voltage-gated ion channels are activated during threshold depolarization, which does not occur during the propagation of a passive electrotonic potential. The process of propagation of the action potential depends on the passive (capacitance, resistance) and active (activation of voltage-controlled channels) properties of the nerve fiber membrane.
Both the internal and external environment of the axon is a good conductor. The axon membrane, despite its insulating properties, can also conduct current due to the presence of ion “leakage” channels. When an unmyelinated fiber is irritated, voltage-gated sodium channels open at the site of irritation, which causes the appearance of an incoming current and the generation of a depolarization phase of the action potential in this section of the axon. The incoming Na + current induces local current circles between the depolarized and non-depolarized regions of the membrane. Due to the described mechanism in unmyelinated fiber, the action potential propagates in both directions from the site of excitation.
In a myelinated nerve fiber, action potentials are generated only at the nodes of Ranvier. The electrical resistance of the areas covered with the myelin sheath is high and does not allow the development of local circular currents, which are necessary to generate an action potential. With the spread of excitation along the myelinated fiber, the nerve impulse jumps from one intercept of Ranvier to another (saltatory conduction) (see Fig. 1.20). In this case, the action potential can spread in both directions from the site of irritation, as in an unmyelinated fiber. Saltatory conduction
 Rice. 1.20. Scheme of the distribution of electrical potential along the nerve fiber.
Rice. 1.20. Scheme of the distribution of electrical potential along the nerve fiber.
A- Propagation of the action potential along the unmyelinated axon: a - axon at rest; b - initiation of the action potential and the occurrence of local currents; c - distribution of local currents; d - propagation of the action potential along the axon. B- Propagation of the action potential from the body of the neuron to the terminal ending. B- Saltatory impulse conduction along the myelinated fiber. Nodes of Ranvier separate segments of the axon myelin sheath
impulse conduction provides a 5-50 times higher speed of excitation compared to unmyelinated fiber. In addition, it is more economical, since local depolarization of the axon membrane only at the node of Ranvier leads to the loss of 100 times less ions than the formation of local currents in an unmyelinated fiber. In addition, during saltatory conduction, voltage-gated potassium channels are minimally involved, as a result of which the action potentials of myelinated fibers often do not have a trace hyperpolarization phase.
Laws of conduction of excitation along the nerve fiber First law: when the nerve fiber is irritated, the excitation spreads along the nerve in both directions.
Second law: propagation of excitation in both directions occurs at the same speed.
Third law: excitation spreads along the nerve without the phenomenon of attenuation, or without decrement. Fourth Law: conduction of excitation along the nerve fiber is possible only with its anatomical and physiological integrity. Any injury to the surface membrane of the nerve fiber (cutting, compression due to inflammation and swelling of the surrounding tissues) disrupts the conduction of irritation. Conduction is also disturbed when the physiological state of the fiber changes: blockade of ion channels, cooling, etc.
Fifth law: excitation of propagation along nerve fibers is isolated, i.e. does not pass from one fiber to another, but excites only those cells with which the endings of this nerve fiber come into contact. Due to the fact that the composition of the peripheral nerve usually includes many different fibers (motor, sensory, vegetative), innervating different organs and tissues and performing different functions, isolated conduction along each fiber is of particular importance.
Sixth law: the nerve fiber does not get tired; the action potential of the fiber has the same amplitude for a very long time.
Seventh Law: the speed of excitation conduction is different in different nerve fibers and is determined by the electrical resistance of the intra- and extracellular environment, the axon membrane, and also by the diameter of the nerve fiber. With an increase in fiber diameter, the rate of conduction of stimulation increases.
Classification of nerve fibers
Based on the speed of conduction of excitation along the nerve fibers, the duration of the phases of the action potential and structural features, three main types of nerve fibers are distinguished: A, B and C.
All type A fibers are myelinated; they are divided into 4 subgroups: α, β, γ and δ. The largest diameter of the αA-fibers (12-22 microns), which determines the high speed of excitation through them (70-170 m/s). αA type fibers in humans conduct excitation from the motor neurons of the anterior horns of the spinal cord to skeletal muscles, as well as from the proprioceptive receptors of muscles to the sensory centers of the central nervous system.
Other fibers type A(β, γ and δ) have a smaller diameter, a slower conduction rate and a longer action potential. These groups of fibers include predominantly sensitive fibers that conduct impulses from various receptors in the central nervous system; the exception is γA fibers, which conduct excitation from γ-neurons of the anterior horns of the spinal cord to intrafusal muscle fibers.
fibers type B also myelinated, related mainly to the preganglionic fibers of the autonomic nervous system. The speed of conduction along them is 3-18 m / s, the duration of the action potential is almost 3 times higher than that of type A fibers. The phase of trace depolarization is not characteristic of these fibers.
fibers type C unmyelinated, have a small diameter (about 1 micron) and a low speed of excitation (up to 3 m/s). Most type C fibers are postganglionic fibers of the sympathetic nervous system, some type C fibers are involved in the conduction of excitation from pain, temperature and other receptors.
1.5. Coding
Information transmitted along the axon in one way or another is encoded. A set of neurons that provide a specific function (for example, a specific sensory modality) forms a projection path (the first encoding method). Thus, the visual pathway includes retinal neurons, the lateral geniculate body of the thalamus, and the visual areas of the cerebral cortex. Axons that conduct visual signals are part of the optic nerve, optic tract, visual radiation. The physiological stimulus for the activation of the visual system is the light that hits the retina. Retinal neurons convert this information and transmit the signal further along the visual pathway. However, with mechanical or electrical stimulation of the neurons of the visual pathway, a visual sensation also arises, although, as a rule, a distorted one. So, the neurons of the visual system make up the projection pathway, upon activation of which a visual sensation arises. Motor pathways also represent projection structures. For example, when certain neurons of the cerebral cortex are activated, discharges are generated in the motor neurons of the muscles of the hand, and these muscles contract.
The second encoding method is due to the principle of ordered spatial (somatotopic) organization of the CNS. Somatotopic maps are compiled by certain groups of neurons in the sensory and motor systems. These groups of neurons, firstly, receive information from appropriately localized areas of the body surface and, secondly, send motor commands to certain parts of the body. In the visual system, areas of the retina are represented in the cerebral cortex by groups of neurons that form retinotopic maps. In the auditory system, the frequency characteristics of sounds are reflected in tonotopic maps.
The third method of encoding information is based on varying the characteristics of sequences (series) of nerve impulses directed to
resulting from synaptic transmission to the next group of neurons, while the coding mechanism is the temporary organization of the discharge of nerve impulses. Possible different types such coding. The average firing rate is often used as a code: in many sensory systems, an increase in stimulus intensity is accompanied by an increase in the firing rate of sensory neurons. In addition, the duration of the discharge, various grouping of pulses in the discharge, the duration of high-frequency bursts of pulses, etc. can serve as a code.
1.6. Carrying out excitation between cells.
Interconnections between nerve cells are carried out by interneuronal contacts, or synapses. Information in the form of a series of action potentials comes from the first (presynaptic) neuron to the second (postsynaptic) either by forming a local current between neighboring cells (electrical synapses), or indirectly by chemicals - mediators, neurotransmitters (chemical synapses), or using both mechanisms ( mixed synapses). Fast signal transmission is carried out by electrical synapses, slower - chemical.
Typical synapses are formations formed by the axon terminals of one neuron and the dendrites of another (axodendritic synapses). In addition, there are axosomatic, axo-axonal and dendrodendritic synapses (Fig. 1.21). Some associative neurons have a variety of synaptic connections (Fig. 1.22). The synapse between a motor neuron axon and a skeletal muscle fiber is called a motor end plate, or neuromuscular junction.
At electrical synapse(Fig. 1.23) the cell membranes of neighboring neurons are closely adjacent to each other, the gap between them is about 2 nm. The sections of the membranes of neighboring cells that form a gap contact contain specific protein complexes consisting of 6 subunits (connexons) arranged in such an order that they form a water-filled pore in the center of the contact. The connexons of the membranes of neighboring cells, lining up against each other, form an open connection - "channels", the distance between which is about 8 nm.
 Rice. 1.21. The main types of synapses.
Rice. 1.21. The main types of synapses.
BUT- a - electrical synapse; b - spiny synapse containing electron-dense vesicles; in - en passant"-synapse, or synaptic "kidney"; d - inhibitory synapse located on the initial part of the axon (contains ellipsoid vesicles); e - dendritic spine; e - spiny synapse; g - inhibitory synapse; h - axo-axonal synapse; and - reciprocal synapse; k - excitatory synapse. B- Atypical synapses: 1 - axo-axonal synapse. The end of one axon can regulate the activity of another; 2 - dendrodendritic synapse; 3 - somasomatic synapse
Electrical synapses are most often formed in the embryonic stage of development, in an adult their number decreases. However, in an adult organism, the significance of electrical synapses is preserved for glial cells and amacrine cells of the retina; electrical synapses can be found in the brain stem, especially in the lower olives, in the retina, vestibular roots.
Depolarization of the presynaptic membrane leads to the formation of a potential difference with the non-depolarized postsynaptic membrane. As a result, through the channels formed by the connexons, the movement of positive ions along the potential difference gradient into the postsynaptic cell or the movement of anions in the opposite direction begins. When reaching on the postsynaptic membrane
 Rice. 1.22. Associative neuron with multiple synaptic connections.
Rice. 1.22. Associative neuron with multiple synaptic connections.
1 - axon hillock, passing into the axon; 2 - myelin sheath; 3 - axodendritic synapse; 4 - core; 5 - dendrite; 6 - axosomatic synapse
 Rice. 1.23. The structure of the electrical synapse.
Rice. 1.23. The structure of the electrical synapse.
BUT- Gap contact between sections of membranes of neighboring cells. B- Connexons of the membranes of neighboring cells form an interneuronal "channel". 1 - protein complex; 2 - ion channel. 3 - channel; 4 - connexon of cell 1; 5 - every six subunits; 6 - cell connexon 2
total depolarization of the threshold value, an action potential arises. It is important to note that in an electrical synapse, ion currents arise with a minimum time delay of 10 -5 s, which explains the high synchronization of the response of even a very large number of cells connected by a gap junction. Conduction of current through an electrical synapse is also possible in both directions (as opposed to a chemical synapse).
The functional state of electrical synapses is regulated by Ca 2+ ions and the level of cell membrane potential, which creates conditions for influencing the propagation of excitation up to its termination. The features of the activity of electrical synapses include the impossibility of direct transfer of excitation to distant cells, since only a few others are directly connected with the excited cell; the level of excitation in presynaptic and postsynaptic cells is the same; slow down the spread
excitation is impossible, in connection with this, the brain of newborns and young children, which contains significantly more electrical synapses than the brain of an adult, turns out to be much more excitable for electrical processes: a rapidly spreading electrical excitation is not subject to inhibitory correction and almost instantly becomes generalized, which explains its special vulnerability and susceptibility to the development of paroxysmal activity.
It should be noted that in some forms of demyelinating polyneuropathies, the axons that are part of one nerve trunk begin to come into close contact with each other, forming pathological zones (ephaps), within which it becomes possible to "jump" the action potential from one axon to another. As a result, symptoms may appear, reflecting the receipt of "pseudo-information" in the brain - a sensation of pain without irritation of peripheral pain receptors, etc.
chemical synapse also transmits an electrical signal from the presynaptic to the postsynaptic cell, but in it, ion channels on the postsynaptic membrane open or close with the help of chemical carriers (mediators, neurotransmitters) released from the presynaptic membrane (Fig. 1.24). Changing the ability to conduct certain ions through the postsynaptic membrane is the basis for the functioning of chemical synapses. Ionic currents change the potential of the postsynaptic membrane, i.e. cause the development of postsynaptic potential. Depending on the conductivity of which ions changes under the action of a neurotransmitter, its effect can be inhibitory (hyperpolarization of the postsynaptic membrane due to an additional outward current of K+ ions or an incoming current of C1 - ions) or excitatory (depolarization of the postsynaptic membrane with an additional incoming current of Ca 2+ ions). or Na+).
In the synapse (Fig. 1.25), a presynaptic process containing presynaptic vesicles (vesicles) and a postsynaptic part (dendrite, cell body or axon) are isolated. In the presynaptic nerve ending, neurotransmitters accumulate in vesicles. Synaptic vesicles are fixed mainly on the cytoskeleton by means of synapsin proteins localized on the cytoplasmic surface of each vesicle and spectrin located on the F-actin fibers of the cytoskeleton (Fig. 1.26). A smaller part of the vesicles is associated with presi-
naptic membrane via the vesicle protein synaptobrevin and the presynaptic membrane protein syntaxin.
One vesicle contains 6000-8000 transmitter molecules, which is 1 transmitter quantum, i.e. the minimum amount released into the synaptic cleft. When a series of action potentials reaches the nerve ending (presynaptic membrane), Ca 2+ ions rush into the cell. On vesicles associated with the presynaptic membrane, Ca 2+ ions bind to the protein of synaptotagmi vesicles.
 Rice. 1.24. The main stages of transmission through a chemical synapse: 1 - the action potential reaches the presynaptic ending; 2 - depolarization of the presynaptic membrane leads to the opening of voltage-dependent Ca 2+ channels; 3 - Ca 2+ ions mediate the fusion of vesicles with the presynaptic membrane; 4 - mediator molecules are released into the synaptic cleft by exocytosis; 5 - mediator molecules bind to postsynaptic receptors, activating ion channels; 6 - there is a change in the conductivity of the membrane for ions and, depending on the properties of the mediator, an excitatory (depolarization) or inhibitory (hyperpolarization) potential of the postsynaptic membrane arises; 7 - ion current propagates along the postsynaptic membrane; 8 - mediator molecules return to the presynaptic ending by reuptake or 9 - diffuse into the extracellular fluid
Rice. 1.24. The main stages of transmission through a chemical synapse: 1 - the action potential reaches the presynaptic ending; 2 - depolarization of the presynaptic membrane leads to the opening of voltage-dependent Ca 2+ channels; 3 - Ca 2+ ions mediate the fusion of vesicles with the presynaptic membrane; 4 - mediator molecules are released into the synaptic cleft by exocytosis; 5 - mediator molecules bind to postsynaptic receptors, activating ion channels; 6 - there is a change in the conductivity of the membrane for ions and, depending on the properties of the mediator, an excitatory (depolarization) or inhibitory (hyperpolarization) potential of the postsynaptic membrane arises; 7 - ion current propagates along the postsynaptic membrane; 8 - mediator molecules return to the presynaptic ending by reuptake or 9 - diffuse into the extracellular fluid
nom, which causes the opening of the vesicle membrane (see Fig. 1.26). In parallel with this, the synaptophysin polypeptide complex fuses with unidentified proteins of the presynaptic membrane, which leads to the formation of a pore through which regulated exocytosis occurs, i.e. secretion of a neurotransmitter into the synaptic cleft. Special vesicle proteins (rab3A) regulate this process.
Ca 2+ ions in the presynaptic terminal activate Ca 2+ -calmodulin-dependent protein kinase II, an enzyme that phosphorylates synapsin on the presynaptic membrane. As a result, transmitter-loaded vesicles can be released from the cytoskeleton and move to the presynaptic membrane for the next cycle.
The width of the synaptic cleft is about 20-50 nm. Neurotransmitter molecules are released into it, the local concentration of which immediately after the release is quite high and is in the millimolar range. Neurotransmitter molecules diffuse to the postsynaptic membrane in about 0.1 ms.
In the postsynaptic membrane, the subsynaptic zone is isolated - the area of \u200b\u200bdirect contact between the presynaptic and postsynaptic membranes, also called the active zone of the synapse. It contains proteins that form ion channels. At rest, these channels rarely open. When neurotransmitter molecules hit the postsynaptic membrane, they interact with ion channel proteins (synaptic receptors), changing their conformation and leading to a much more frequent opening of ion channels. Those receptors whose ion channels open upon direct contact with a ligand (neurotransmitter) are called ionotropic. Receptors in which open
 Rice. 1.25. Ultrastructure of the axodendritic synapse. 1 - axon; 2 - dendrite; 3 - mitochondria; 4 - synaptic vesicles; 5 - presynaptic membrane; 6 - postsynaptic membrane; 7 - synaptic cleft
Rice. 1.25. Ultrastructure of the axodendritic synapse. 1 - axon; 2 - dendrite; 3 - mitochondria; 4 - synaptic vesicles; 5 - presynaptic membrane; 6 - postsynaptic membrane; 7 - synaptic cleft
The development of ion channels is associated with the connection of other chemical processes, called metabotropic(Fig. 1.27).
In many synapses, neurotransmitter receptors are located not only on the postsynaptic but also on the presynaptic membrane. (autoreceptors). When a neurotransmitter interacts with the autoreceptors of the presynaptic membrane, its release is enhanced or weakened (positive or negative feedback), depending on the type of synapse. The functional state of autoreceptors is also affected by the concentration of Ca 2+ ions.
Interacting with the postsynaptic receptor, the neurotransmitter opens nonspecific ion channels in the postsynaptic
 Rice. 1.26. Vesicle docking at the presynaptic membrane. BUT- The synaptic vesicle is attached to the cytoskeletal element with the help of a synapsin molecule. The docking complex is highlighted by a quadrilateral: 1 - Samkinase 2; 2 - synapsis 1; 3 - fodrin; 4 - mediator carrier; 5 - synaptophysin; 6 - docking complex
Rice. 1.26. Vesicle docking at the presynaptic membrane. BUT- The synaptic vesicle is attached to the cytoskeletal element with the help of a synapsin molecule. The docking complex is highlighted by a quadrilateral: 1 - Samkinase 2; 2 - synapsis 1; 3 - fodrin; 4 - mediator carrier; 5 - synaptophysin; 6 - docking complex
B- An enlarged scheme of the docking complex: 7 - synaptobrevin; 8 - synaptotagmin; 9 - rab3A; 10-NSF; 11 - synaptophysin; 12 - SNAP; 13 - syntaxin; 14 - neurexin; 15 - fizofillin; 16 - α-SNAP; 17 - Ca 2+; 18 - n-sec1. CaM kinase-2 - calmodulin-dependent protein kinase 2; n-secl - secretory protein; NSF - N-ethylmaleimide-sensitive fusion protein; gab33A - GTPase from the ras family; SNAP - presynaptic membrane protein
membrane. Excitatory postsynaptic potential arises due to an increase in the ability of ion channels to conduct monovalent cations, depending on their electrochemical gradients. Thus, the potential of the postsynaptic membrane is in the range between -60 and -80 mV. The equilibrium potential for Na+ ions is +55 mV, which explains the strong driving force for Na+ ions into the cell. The equilibrium potential for K+ ions is approximately -90 mV, i.e. a slight current of K+ ions remains, directed from the intracellular environment to the extracellular one. The work of ion channels leads to depolarization of the postsynaptic membrane, which is called the excitatory postsynaptic potential. Since ion currents depend on the difference between the equilibrium potential and the membrane potential, then at a reduced resting potential of the membrane, the current of Na + ions weakens, and the current of K + ions increases, which leads to a decrease in the amplitude of the excitatory postsynaptic potential. Na + and K + currents involved in the occurrence of excitatory postsynaptic
 Rice. 1.27. Diagram of the structure of the receptor.
Rice. 1.27. Diagram of the structure of the receptor.
BUT- Metabotropic. B- Ionotropic: 1 - neuromodulators or drugs; 2 - receptors with different binding sites (heteroceptor); 3 - neuromodulation; 4 - secondary messenger; 5 - autoreceptor; 6 - feedback; 7 - embedding of the vesicle membrane; 8 - neuromodulators; 9 - transmitter; 10 - neuromodulation; 11-transmitter catalyzes the reactions of G-proteins; 12 - the transmitter opens the ion channel
which potential, behave differently than when generating an action potential, since other ion channels with different properties take part in the mechanism of postsynaptic depolarization. If, during the generation of an action potential, voltage-gated ion channels are activated, and with an increase in depolarization, other channels open, as a result of which the depolarization process strengthens itself, then the conductivity of transmitter-gated (ligand-gated) channels depends only on the number of transmitter molecules bound to the receptors, i.e. on the number of open ion channels. The amplitude of the excitatory postsynaptic potential ranges from 100 μV to 10 mV, the duration of the potential is in the range from 4 to 100 ms, depending on the type of synapse.
An excitatory postsynaptic potential formed locally in the synapse zone passively propagates throughout the entire postsynaptic membrane of the cell. With the simultaneous excitation of a large number of synapses, the phenomenon of summation of the postsynaptic potential occurs, manifested by a sharp increase in its amplitude, as a result of which the membrane of the entire postsynaptic cell can be depolarized. If the amount of depolarization reaches the threshold value (more than 10 mV), then the generation of an action potential begins, which is carried out along the axon of the postsynaptic neuron. About 0.3 ms elapses from the beginning of the excitatory postsynaptic potential to the formation of the action potential, i.e. with a massive release of the neurotransmitter, the postsynaptic potential may appear already after 0.5-0.6 ms from the moment the action potential arrives in the presynaptic region (the so-called synaptic delay).
Other compounds may have a high affinity for the postsynaptic receptor protein. Depending on what (in relation to the neurotransmitter) effect their binding to the receptor leads to, agonists (unidirectional action with the neurotransmitter) and antagonists (the action of which prevents the effects of the neurotransmitter) are isolated.
There are receptor proteins that are not ion channels. When neurotransmitter molecules bind to them, a cascade of chemical reactions occurs, as a result of which neighboring ion channels open with the help of secondary messengers - metabotropic receptors. G-protein plays an important role in their functioning. Synaptic transmission, which uses metabotropic reception, is very slow, with an impulse transmission time of about 100 ms. to the synapses
this type includes postganglionic receptors, receptors of the parasympathetic nervous system, autoreceptors. An example is the muscarinic-type cholinergic synapse, in which the neurotransmitter binding site and ion channel are not localized in the transmembrane protein itself, the metabotropic receptors are directly coupled to the G protein. When the transmitter binds to the receptor, the G protein, which has three subunits, forms a complex with the receptor. GDP bound to the G-protein is replaced by GTP, while the G-protein is activated and acquires the ability to open the potassium ion channel, i.e. hyperpolarize the postsynaptic membrane (see Fig. 1.27).
Second messengers can open or close ion channels. Thus, ion channels can be opened by cAMP/IP 3 or phosphorylation of protein kinase C. This process also occurs with the help of G-protein, which activates phospholipase C, which leads to the formation of inositol triphosphate (IP 3). Additionally, the formation of diacylglycerol (DAG) and protein kinase C (PKC) increases (Fig. 1.28).
Each nerve cell has on its surface many synaptic endings, some of which are excitatory, others are torsion.
 Rice. 1.28. Role of second messengers of inositol triphosphate (IP 3) (BUT) and diacylglycerol (DAG) (B) in the work of the metabotropic receptor. When the mediator binds to the receptor (P), the conformation of the G-protein changes, followed by the activation of phospholipase C (PLC). Activated FLS cleaves phosphatidylinositol triphosphate (PIP 2 ) into DAG and IP 3 . DAG remains in the inner layer of the cell membrane, and IP 3 diffuses into the cytosol as a second messenger. DAG is embedded in the inner layer of the membrane where it interacts with protein kinase C (PKC) in the presence of phosphatidylserine (PS)
Rice. 1.28. Role of second messengers of inositol triphosphate (IP 3) (BUT) and diacylglycerol (DAG) (B) in the work of the metabotropic receptor. When the mediator binds to the receptor (P), the conformation of the G-protein changes, followed by the activation of phospholipase C (PLC). Activated FLS cleaves phosphatidylinositol triphosphate (PIP 2 ) into DAG and IP 3 . DAG remains in the inner layer of the cell membrane, and IP 3 diffuses into the cytosol as a second messenger. DAG is embedded in the inner layer of the membrane where it interacts with protein kinase C (PKC) in the presence of phosphatidylserine (PS)
muscular. If adjacent excitatory and inhibitory synapses are activated in parallel, the resulting currents are superimposed on each other, resulting in a postsynaptic potential with an amplitude smaller than its excitatory and inhibitory components separately. At the same time, hyperpolarization of the membrane is significant due to an increase in its conductivity for K + and C1 - ions.
Thus, an excitatory postsynaptic potential is generated due to an increase in Na+ ion permeability and an incoming Na+ ion current, and an inhibitory postsynaptic potential is generated due to an outgoing K+ ion current or an incoming C1 - ion current. A decrease in conductivity for K + ions should depolarize the cell membrane. Synapses, in which depolarization is caused by a decrease in conductivity for K + ions, are localized in the ganglia of the autonomic (autonomous) nervous system
Synaptic transfer must be completed quickly so that the synapse is ready for a new transfer, otherwise the response would not arise under the influence of newly incoming signals, would be observed depolarization block. An important regulatory mechanism is a rapid decrease in the sensitivity of the postsynaptic receptor (desensitization), which occurs when the neurotransmitter molecules are still preserved. Despite the continuous binding of the neurotransmitter to the receptor, the conformation of the channel-forming protein changes, the ion channel becomes impermeable to ions, and the synaptic flow stops. In many synapses, receptor desensitization can be prolonged (up to several minutes) until the channel is reconfigured and reactivated.
Other ways to terminate the action of the transmitter, which avoid long-term desensitization of the receptor, are the rapid chemical cleavage of the transmitter into inactive components or its removal from the synaptic cleft by highly selective reuptake by the presynaptic ending. The nature of the inactivating mechanism depends on the type of synapse. So, acetylcholine is very quickly hydrolyzed by acetylcholinesterase into acetate and choline. In the CNS, excitatory glutamatergic synapses are densely covered with processes of astrocytes that actively capture the neurotransmitter from the synaptic cleft and metabolize it.
1.7. Neurotransmitters and neuromodulators
Neurotransmitters transmit a signal in synapses between neurons or between neurons and executive organs (muscle, glandular cells). Neuromodulators presynaptically affect the amount of neurotransmitter released or its reuptake by the neuron. In addition, neuromodulators postsynaptically regulate the sensitivity of receptors. Thus, neuromodulators are able to regulate the level of excitability in synapses and change the effect of neurotransmitters. Neurotransmitters and neuromodulators together form a group of neuroactive substances.
Many neurons are targeted by several neuroactive substances but release only one transmitter when stimulated. The same neurotransmitter, depending on the type of postsynaptic receptor, can have an excitatory or inhibitory effect. Some neurotransmitters (such as dopamine) can also function as neuromodulators. A neurofunctional system usually involves several neuroactive substances, and one neuroactive substance is able to influence several neurofunctional systems.
Catecholaminergic neurons
Catecholaminergic neurons contain neurotransmitters such as dopamine, norepinephrine, or epinephrine in the perikarya and processes, which are synthesized from the amino acid tyrosine. In the adult brain, dopaminergic, noradrenergic, and adrenergic neurons correspond in localization to melanin-containing neurons. Noradrenergic and dopaminergic cells are numbered from A1 to A15, and adrenergic cells are numbered from C1 to C3, sequence numbers assigned in ascending order, according to the location in the brainstem from the lower to the upper sections.
dopaminergic neurons Dopamine-synthesizing cells (A8-A15) are located in the midbrain, diencephalon, and telencephalon (Fig. 1.29). The largest group of dopaminergic cells is the compact part of the substantia nigra (A9). Their axons form an ascending path passing through the lateral part of the hypothalamus and the internal capsule, nigrostriatal hair bundles
 Rice. 1.29. Localization of dopaminergic neurons and their pathways in the rat brain.
Rice. 1.29. Localization of dopaminergic neurons and their pathways in the rat brain.
1 - cerebellum; 2 - cerebral cortex; 3 - striatum; 4 - nucleus accumbens; 5 - frontal cortex; 6 - olfactory bulb; 7 - olfactory tubercle; 8 - caudate nucleus; 9 - almond-shaped nucleus; 10 - median elevation; 11 - nigrostriatal bundle. The main pathway (nigrostriatal bundle) starts in the substantia nigra (A8, A9) and runs forward to the striatum
con reach the caudate nucleus and the shell. Together with dopaminergic neurons of the reticular substance (A8), they form the nigrostriatal system.
The main pathway (nigrostriatal bundle) originates in the substantia nigra (A8, A9) and runs forward to the striatum.
The mesolimbic group of dopaminergic neurons (A10) extends from the mesencephalic regions to the limbic system. Group A10 forms the ventral apex at the interpeduncular nuclei in the tegmentum of the midbrain. Axons go to the internal nuclei of the terminal sulcus, septum, olfactory tubercles, nucleus accumbens (n. accumbens), cingulate gyrus.
The third dopaminergic system (A12), called tuberoinfundibular, is located in the diencephalon, is located in the gray tubercle and extends to the infundibulum. This system is associated with neuroendocrine functions. Other diencephalic groups of cells (A11, A13 and A14) and their target cells are also located in the hypothalamus. A small group of A15 is dispersed in the olfactory bulb and is the only dopaminergic group of neurons in the telencephalon.
All dopamine receptors act through the second messenger system. Their postsynaptic action can be excitatory or inhibitory. Dopamine is rapidly taken back to the presynaptic terminal where it is metabolized by monoamine oxidase (MAO) and catechol-O-methyltransferase (COMT).
Noradrenergic neurons Noradrenergic nerve cells are located only in a narrow anterolateral zone of the tegmentum of the medulla oblongata and pons (Fig. 1.30). In-
 Rice. 1.30. Localization of noradrenergic neurons and their pathways in the rat brain (parasagittal section).
Rice. 1.30. Localization of noradrenergic neurons and their pathways in the rat brain (parasagittal section).
1 - cerebellum; 2 - dorsal bundle; 3 - ventral bundle; 4 - hippocampus; 5 - cerebral cortex; 6 - olfactory bulb; 7 - partition; 8 - medial anterior cerebral bundle; 9 - end strip; 10 - hypothalamus.
The main pathway begins in the locus coeruleus (A6) and runs forward in several bundles, giving offshoots to various parts of the brain. Also, noradrenergic nuclei are located in the ventral part of the brainstem (A1, A2, A5 and A7). Most of their fibers go along with the fibers of neurons in the coeruleus, however, some are projected in the dorsal direction.
the filaments coming from these neurons ascend to the midbrain or descend to the spinal cord. In addition, noradrenergic cells have connections with the cerebellum. Noradrenergic fibers branch more extensively than dopaminergic ones. They are believed to play a role in the regulation of cerebral blood flow.
The largest group of noradrenergic cells (A6) is located in the locus coeruleus (locus cereleus) and includes almost half of all noradrenergic cells (Fig. 1.31). The nucleus is located in the upper part of the bridge at the bottom of the IV ventricle and extends up to the inferior colliculi of the quadrigemina. The axons of the cells of the blue spot branch many times, their adrenergic endings can be found in many parts of the CNS. They have a modulating effect on the processes of maturation and learning, information processing in the brain, sleep regulation, and endogenous pain inhibition.
The posterior noradrenergic bundle originates from the A6 group and connects in the midbrain with the nuclei of the posterior suture, the superior and inferior tubercles of the quadrigemina; in the diencephalon - with the anterior nuclei of the thalamus, medial and lateral geniculate bodies; in the final brain - with the amygdala, hippocampus, neocortex, cingulate gyrus.
Additional fibers from the cells of the A6 group go to the cerebellum through its superior peduncle (see Fig. 1.31). Descending fibers from the locus coeruleus, together with the fibers of the neighboring group of A7 cells, go to the posterior nucleus of the vagus nerve, the inferior olive and the spinal cord. Anterolateral-
 Rice. 1.31. Scheme of conducting noradrenergic pathways from the blue nucleus (spot) located in the gray matter of the bridge.
Rice. 1.31. Scheme of conducting noradrenergic pathways from the blue nucleus (spot) located in the gray matter of the bridge.
1 - fibers of the conductive path; 2 - hippocampus; 3 - thalamus; 4 - hypothalamus and amygdala; 5 - cerebellum; 6 - spinal cord; 7 - blue spot
The descending bundle from the locus coeruleus sends fibers to the anterior and posterior horns of the spinal cord.
The neurons of groups A1 and A2 are located in the medulla oblongata. Together with groups of pontine cells (A5 and A7), they form the anterior ascending noradrenergic pathways. In the midbrain, they are projected onto the gray periaqueductal nucleus and the reticular formation, in the diencephalon - onto the entire hypothalamus, and in the telencephalon - onto the olfactory bulb. In addition, bulbospinal fibers also go from these groups of cells (A1, A2, A5, A7) to the spinal cord.
In the PNS, norepinephrine (and to a lesser extent epinephrine) is an important neurotransmitter of the sympathetic postganglionic endings of the autonomic nervous system.
Adrenergic neurons
Adrenaline-synthesizing neurons are located only in the medulla oblongata, in a narrow anterolateral region. The largest group of C1 cells lies behind the posterior olive nucleus, the middle group of C2 cells - next to the nucleus of the solitary pathway, the group of C3 cells - directly under the periaqueductal gray matter. Efferent pathways from C1-C3 go to the posterior nucleus of the vagus nerve, the nucleus of the solitary tract, the blue spot, the periaqueductal gray matter of the pons and midbrain, and the hypothalamus.
There are 4 main types of catecholaminergic receptors, differing in response to the action of agonists or antagonists and in postsynaptic effects. α1 receptors control calcium channels via the second messenger inositol phosphate-3 and, when activated, increase intracellular ion concentration
Ca 2+ . Stimulation of β2 receptors leads to a decrease in the concentration of the second messenger cAMP, which is accompanied by various effects. B receptors, through the secondary messenger cAMP, increase membrane conductivity for K+ ions, generating an inhibitory postsynaptic potential.
Serotonergic neurons
Serotonin (5-hydroxytryptamine) is formed from the amino acid tryptophan. Most serotonergic neurons are localized in the medial parts of the brainstem, forming the so-called raphe nuclei (Fig. 1.32). Groups B1 and B2 are located in the medulla oblongata, B3 - in the border zone between the medulla oblongata and the bridge, B5 - in the bridge, B7 - in the midbrain. The raphe neurons B6 and B8 are located in the tegmentum pons and the midbrain. The raphe nuclei also contain nerve cells containing other neurotransmitters such as dopamine, norepinephrine, GABA, enkephalin, and substance P. For this reason, the raphe nuclei are also called multitransmitter centers.
Projections of serotonergic neurons correspond to the course of norepinephrine fibers. The bulk of the fibers goes to the structures of the limbic system, the reticular formation and the spinal cord. There is a connection with the blue spot - the main concentration of norepinephrine neurons.
The large anterior ascending tract ascends from cells of the B6, B7, and B8 groups. It goes anteriorly through the midbrain tegmentum and laterally through the hypothalamus, then gives off branches towards the fornix and cingulate gyrus. Through this pathway, groups B6, B7 and B8 are connected in the midbrain with the interpeduncular nuclei and substantia nigra, in the diencephalon with the nuclei of the leash, thalamus and hypothalamus, in the final brain with the nuclei of the septum and the olfactory bulb.
There are numerous projections of serotonergic neurons to the hypothalamus, cingulate gyrus, and olfactory cortex, as well as connections to the striatum and frontal cortex. The shorter posterior ascending tract connects the cells of groups B3, B5, and B7 via the posterior longitudinal bundle to the periaqueductal gray matter and the posterior hypothalamic region. In addition, there are serotonergic projections to the cerebellum (from B6 and B7) and the spinal cord (from B1 to B3), as well as numerous fibers connecting to the reticular formation.
The release of serotonin occurs in the usual way. Receptors are located on the postsynaptic membrane, which, with the help of secondary messengers, open channels for K + and Ca 2+ ions. There are 7 classes of serotonin receptors: 5-HT 1 - 5-HT 7 that respond differently to the action of agonists and antagonists. Receptors 5-HT 1 , 5-HT 2 and 5-HT 4 are located in the brain, receptors 5-HT 3 - in the PNS. The action of serotonin is terminated by the mechanism of neurotransmitter reuptake by the presynaptic ending. Serotonin that has not entered the vesicles is deaminated with the help of MAO. There is an inhibitory effect of descending serotonergic fibers on the first sympathetic neurons of the spinal cord. It is assumed that in this way the raphe neurons of the medulla oblongata control the conduction of pain impulses in the anterolateral system. Serotonin deficiency is associated with depression.
 Rice. 1.32. Localization of serotonergic neurons and their pathways in the rat brain (parasagittal section).
Rice. 1.32. Localization of serotonergic neurons and their pathways in the rat brain (parasagittal section).
1 - olfactory bulb; 2 - belt; 3 - corpus callosum; 4 - cerebral cortex; 5 - medial longitudinal bundle; 6 - cerebellum; 7 - medial anterior cerebral bundle; 8 - brain strip; 9 - end strip; 10 - vault; 11 - caudate nucleus; 12 - outer capsule. Serotonergic neurons are grouped into nine nuclei located in the brainstem. The B6-B9 nuclei project anteriorly to the diencephalon and telencephalon, while the caudal nuclei project to the medulla oblongata and spinal cord.
Histaminergic neurons
Histaminergic nerve cells are located in the lower part of the hypothalamus close to the infundibulum. Histamine is metabolized by the enzyme histidine decarboxylase from the amino acid histidine. Long and short bundles of fibers of histaminergic nerve cells in the lower part of the hypothalamus go to the brain stem as part of the posterior and periventricular zones. Histaminergic fibers reach the periaqueductal gray matter, posterior raphe nucleus, medial vestibular nucleus, solitary tract nucleus, posterior vagus nucleus, nucleus
facial nerve, anterior and posterior cochlear nuclei, lateral loop and inferior tubercle of the quadrigemina. In addition, the fibers are sent to the diencephalon - the posterior, lateral and anterior sections of the hypothalamus, mastoid bodies, optic tubercle, periventricular nuclei, lateral geniculate bodies and to the telencephalon - Broca's diagonal gyrus, n. accumbens, amygdala and cerebral cortex.
Cholinergic neurons
Alpha (α)- and gamma (γ)-motoneurons of the oculomotor, trochlear, trigeminal, abducens, facial, glossopharyngeal, vagus, accessory and hypoglossal nerves and spinal nerves are cholinergic (Fig. 1.33). Acetylcholine affects the contraction of skeletal muscles. The preganglionic neurons of the autonomic nervous system are cholinergic, they stimulate the postganglionic neurons of the autonomic nervous system. Other cholinergic nerve cells were given an alphanumeric designation from top to bottom (in reverse order compared to catecholaminergic and serotonergic neurons). Ch1 cholinergic neurons form about 10% of the cells of the median nuclei of the septum, Ch2 neurons make up 70% of the cells of the vertical limb of the diagonal Broca's sulcus, Ch3 neurons make up 1% of the cells of the horizontal limb of the diagonal of Broca's sulcus. All three groups of neurons project downward onto the medial nuclei of the leash and the interpeduncular nuclei. The Ch1 neurons are connected by ascending fibers through the fornix to the hippocampus. The Ch3 cell group is synaptically connected to the nerve cells of the olfactory bulb.
In the human brain, the group of Ch4 cells is relatively extensive and corresponds to the basal nucleus of Meinert, in which 90% of all cells are cholinergic. These nuclei receive afferent impulses from the subcortical diencephalic-telencephalic divisions and form the limbic-paralimbic cortex of the brain. The anterior cells of the nucleus basalis project onto the frontal and parietal neocortex, while the posterior cells project onto the occipital and temporal neocortex. Thus, the nucleus basalis is the transmission link between the limbic-paralimbic regions and the neocortex. Two small groups of cholinergic cells (Ch5 and Ch6) are located in the pons and are considered part of the ascending reticular system.
A small group of cells of the periolivar nucleus, partly consisting of cholinergic cells, is located at the edge of the trapezoid body in the lower parts of the bridge. Its efferent fibers go to the receptor cells of the auditory system. This cholinergic system influences the transmission of sound signals.
Aminacidergic neurons
Neurotransmitter properties have been proven for four amino acids: excitatory for glutamic (glutamate), aspartic (aspartate) acids, and inhibitory for g-aminobutyric acid and glycine. The neurotransmitter properties of cysteine are assumed (excitatory); taurine, serine and p-alanine (brake).
 Rice. 1.33. Localization of cholinergic neurons and their pathways in the rat brain (parasagittal section). 1 - almond-shaped nucleus; 2 - anterior olfactory nucleus; 3 - arcuate core; 4 - basal nucleus of Meinert; 5 - cerebral cortex; 6 - shell of the caudate nucleus; 7 - diagonal Broca's beam; 8 - bent beam (Meinert's beam); 9 - hippocampus; 10 - interpeduncular nucleus; 11 - lateral-dorsal core of the tire; 12 - medial core of the leash; 13 - olfactory bulb; 14 - olfactory tubercle; 15 - reticular formation; 16 - brain strip; 17 - thalamus; 18 - reticular formation of the tire
Rice. 1.33. Localization of cholinergic neurons and their pathways in the rat brain (parasagittal section). 1 - almond-shaped nucleus; 2 - anterior olfactory nucleus; 3 - arcuate core; 4 - basal nucleus of Meinert; 5 - cerebral cortex; 6 - shell of the caudate nucleus; 7 - diagonal Broca's beam; 8 - bent beam (Meinert's beam); 9 - hippocampus; 10 - interpeduncular nucleus; 11 - lateral-dorsal core of the tire; 12 - medial core of the leash; 13 - olfactory bulb; 14 - olfactory tubercle; 15 - reticular formation; 16 - brain strip; 17 - thalamus; 18 - reticular formation of the tire
Glutamatergic and aspartatergic neurons The structurally similar amino acids glutamate and aspartate (Figure 1.34) are classified electrophysiologically as excitatory neurotransmitters. Nerve cells containing glutamate and/or aspartate as neurotransmitters are present in the auditory system (first-order neurons), in the olfactory system (combining the olfactory bulb with the cerebral cortex), in the limbic system, in the neocortex (pyramidal cells). Glutamate is also found in the neurons of the pathways coming from the pyramidal cells: the corticostriate, corticothalamic, corticothectal, corticospinal, and corticospinal tracts.
An important role in the functioning of the glutamate system is played by astrocytes, which are not passive elements of the nervous system, but are involved in providing neurons with energy substrates in response to an increase in synaptic activity. Astrocyte processes -
 Rice. 1.34. Synthesis of glutamic and aspartic acids.
Rice. 1.34. Synthesis of glutamic and aspartic acids.
Glycolysis converts glucose to pyruvate, which, in the presence of acetyl-CoA, enters the Krebs cycle. Further, by transamination, oxaloacetate and α-ketoglutarate are converted into aspartate and glutamate, respectively (reactions are presented at the bottom of the figure)
ki are located around synaptic contacts, which allows them to detect an increase in the synaptic concentration of neurotransmitters (Fig. 1.35). The transport of glutamate from the synaptic cleft is mediated by specific transport systems, two of which are glial-specific ( GLT-1 And GLAST- carriers). Third transport system (EAAC-1), located exclusively in neurons, is not involved in the transfer of glutamate released from synapses. The transition of glutamate into astrocytes occurs along the electrochemical gradient of Na + ions.
Under normal conditions, the relative constancy of extracellular concentrations of glutamate and aspartate is maintained. Their increase includes compensatory mechanisms: capture by neurons and astrocytes of excesses from the intercellular space, presynaptic inhibition of the release of neurotransmitters, metabolic utilization and
 Rice. 1.35. The structure of the glutamatergic synapse.
Rice. 1.35. The structure of the glutamatergic synapse.
Glutamate is released from synaptic vesicles into the synaptic cleft. The figure shows two reuptake mechanisms: 1 - back to the presynaptic ending; 2 - into the neighboring glial cell; 3 - glial cell; 4 - axon; 5 - glutamine; 6 - glutamine synthetase; 7 - ATP + NH 4 +; 8 - glutaminase; 9 - glutamate + NH 4 +; 10 - glutamate; 11 - postsynaptic membrane. In glial cells, glutamine synthase converts glutamate to glutamine, which is then transferred to the presynaptic terminal. At the presynaptic terminal, glutamine is converted back to glutamate by the enzyme glutaminase. Free glutamate is also synthesized in the reactions of the Krebs cycle in mitochondria. Free glutamate is collected in synaptic vesicles until the next action potential occurs. The right side of the figure shows the conversion reactions of glutamate and glutamine mediated by glutamine synthetase and glutaminase
etc. In violation of their elimination from the synaptic cleft, the absolute concentration and residence time of glutamate and aspartate in the synaptic cleft exceed the permissible limits, and the process of depolarization of neuronal membranes becomes irreversible.
There are families of ionotropic and metabotropic glutamate receptors in the mammalian CNS. Ionotropic receptors regulate the permeability of ion channels and are classified according to their sensitivity to the action of N-methyl-D-aspartate (NMDA)α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMRA), kainic acid (K) and L-2-amino-4-phosphonobutyric acid (L-AP4)- the most selective ligands of this type of receptors. The names of these compounds were assigned to the corresponding types of receptors: NMDA, AMPA, K And L-AP4.
The most studied receptors are NMDA-type (Fig. 1.36). postsynaptic receptor NMDA is a complex supramolecular formation that includes several sites (sites) of regulation: a specific binding site for a mediator (L-glutamic acid), a specific binding site for a coagonist (glycine), and allosteric modulatory sites located both on the membrane (polyamine) and in the ion channel , coupled to the receptor (binding sites for divalent cations and "phencyclidine" site - binding site for non-competitive antagonists).
Ionotropic receptors play a key role in the implementation of excitatory neurotransmission in the CNS, the implementation of neuroplasticity, the formation of new synapses (synaptogenesis), and in increasing the efficiency of the functioning of existing synapses. These processes are largely associated with the mechanisms of memory, learning (acquisition of new skills), compensation of functions impaired due to organic damage to the brain.
Excitatory aminoacidergic neurotransmitters (glutamate and aspartate) are cytotoxic under certain conditions. When they interact with overexcited postsynaptic receptors, dendrosomatic lesions develop without changes in the conductive part of the nerve cell. Conditions that create such an overexcitation are characterized by increased release and/or reduced reuptake of the carrier. Overexcitation of glutamate receptors NMDA leads to the opening of ago-
nist-dependent calcium channels and a powerful influx of Ca 2+ into neurons with a sudden increase in its concentration to the threshold. Caused by excess action of aminoacidergic neurotransmitters "excitotoxic death of neurons" is a universal mechanism of damage to the nervous tissue. It underlies the necrotic death of neurons in various brain diseases, both acute (ischemic stroke) and chronic (neuro-
 Rice. 1.36. Glutamate NMDA receptor
Rice. 1.36. Glutamate NMDA receptor
rodegeneration). The extracellular levels of aspartate and glutamate, and hence the severity of excitotoxicity, are affected by the temperature and pH of the brain, extracellular concentrations of monovalent ions C1 - and Na + . Metabolic acidosis inhibits the transport systems of glutamate from the synaptic cleft.
There is evidence of the neurotoxic properties of glutamate associated with the activation of AMPA and K receptors, which leads to a change in the permeability of the postsynaptic membrane for monovalent K+ and Na+ cations, an increase in the incoming current of Na+ ions, and a short-term depolarization of the postsynaptic membrane, which, in turn, causes an increase in influx of Ca 2+ into the cell through agonist-dependent (receptors NMDA) and voltage-gated channels. The flow of Na+ ions is accompanied by the entry of water into the cells, which causes swelling of the apical dendrites and lysis of neurons (osmolytic damage to neurons).
G-protein-coupled metabotropic glutamate receptors play an important role in the regulation of intracellular calcium current caused by the activation of NMDA receptors and perform modulatory functions, thereby causing changes in cell activity. These receptors do not affect the functioning of ion channels, but stimulate the formation of intracellular mediators of diacylglycerol and nositol triphosphate, which are involved in further processes of the ischemic cascade.
GABAergic neurons
Some neurons contain g-aminobutyric acid (GABA) as a neurotransmitter, which is formed from glutamic acid by the action of glutamate decarboxylase (Fig. 1.37). In the cerebral cortex, GABAergic neurons are located in the olfactory and limbic areas (hippocampal basket neurons). GABA also contains neurons of efferent extrapyramidal striatonigral, pallidonigral and subtalamopallidar pathways, cerebellar Purkinje cells, cerebellar cortex neurons (Golgi, stellate and basket), intercalary inhibitory neurons of the spinal cord.
GABA is the most important inhibitory neurotransmitter in the CNS. The main physiological role of GABA is the creation of a stable balance between excitatory and inhibitory systems, modulation and regulation of the activity of the main excitatory neurotransmitter glutamate. GABA limits the spread of an excitatory stimulus both presynaptically - through GABA-B receptors, functionally
 Rice. 1.37. The reaction of the conversion of glutamate to GABA.
Rice. 1.37. The reaction of the conversion of glutamate to GABA.
The coenzyme pyridoxal phosphate is required for the activity of glutamic acid decarboxylase (DHA)
 Rice. 1.38. GABA receptor.
Rice. 1.38. GABA receptor.
1 - benzodiazepine-binding site;
2 - GABA-binding site; 3 - ion channel for CL - ; 4 - barbiturate-binding site
but associated with voltage-dependent calcium channels of presynaptic membranes, and postsynaptically - through GABA-receptors (GABA-barbiturate-benzodiazepine-receptor complex), functionally associated with voltage-dependent chloride channels. Activation of postsynaptic GABA-A receptors leads to hyperpolarization of cell membranes and inhibition of the excitatory impulse caused by depolarization.
The density of GABA-A receptors is maximum in the temporal and frontal cortex, hippocampus, amygdala and hypothalamic nuclei, substantia nigra, periaqueductal gray matter, and cerebellar nuclei. To a lesser extent, receptors are present in the caudate nucleus, putamen, thalamus, occipital cortex, and epiphysis. All three subunits of the GABA-A receptor (α, β and γ) bind GABA, although the binding affinity is highest for the b-subunit (Fig. 1.38). Barbiturates interact with a- and P-subunits; benzodiazepines - only with a 7-subunit. The binding affinity of each of the ligands is increased if other ligands interact in parallel with the receptor.
Glycinergic neurons Glycine is an inhibitory neurotransmitter in almost all parts of the CNS. The highest density of glycine receptors was found in the structures of the brainstem, the cerebral cortex, the striatum, the nuclei of the hypothalamus, the conductors from the frontal cortex to the hypothalamus, and the brain.
gallbladder, spinal cord. Glycine exhibits inhibitory properties through interaction not only with its own strychnine-sensitive glycine receptors, but also with GABA receptors.
In small concentrations, glycine is necessary for the normal functioning of glutamate receptors. NMDA. Glycine is a receptor co-agonist NMDA, since their activation is possible only if glycine binds to specific (insensitive to strychnine) glycine sites. Potentiating effect of glycine on receptors NMDA appears at concentrations below 0.1 µmol, and at concentrations from 10 to 100 µmol, the glycine site is completely saturated. High concentrations of glycine (10-100 mmol) do not activate NMDA-induced depolarization in vivo and therefore do not increase excitotoxicity.
Peptidergic neurons
The neurotransmitter and/or neuromodulatory function of many peptides is still being studied. Peptidergic neurons include:
Hypothalamoneurohypophyseal nerve cells with peptides ok-
Sitocin and vasopressin as neurotransmitters; pituitary cells with peptides somatostatin, corti-
koliberin, thyroliberin, luliberin;
Neurons with peptides of the autonomic nervous system of the gastrointestinal tract, such as substance P, vasoactive intestinal polypeptide (VIN) and cholecystokinin;
Neurons whose peptides are formed from pro-opiomelanocortin (corticotropin and β-endorphin),
Enkephalinergic nerve cells.
Substance-R - containing neurons Substance P is an 11 amino acid peptide that has a slow onset and long-lasting stimulating effect. Substance P contains:
About 1/5 cells of the spinal ganglia and trigeminal (Gasserov) ganglion, the axons of which have a thin myelin sheath or are not myelinated;
Olfactory bulb cells;
Neurons of the periaqueductal gray matter;
Neurons of the path from the midbrain to the interpeduncular nuclei;
Neurons of the efferent nigrostriatal pathways;
Small nerve cells located in the cerebral cortex, mainly in layers V and VI.
VIP-containing neurons Vasoactive intestinal polypeptide (VIP) consists of 28 amino acids. In the nervous system, VIP is an excitatory neurotransmitter and/or neuromodulator. The highest VIP concentration is found in the neocortex, predominantly in bipolar cells. In the brainstem, VIP-containing nerve cells are located in the nucleus of the solitary tract and are associated with the limbic system. The suprachiasmatic nucleus contains VIP-containing neurons associated with the nuclei of the hypothalamus. In the gastrointestinal tract, it has a vasodilating effect and stimulates the transition of glycogen to glucose.
β-Endorphin-containing neuronsβ-Endorphin is a 31 amino acid peptide that functions as an inhibitory neuromodulator in the brain. Endorphinergic cells are found in the mediobasal hypothalamus and in the lower portions of the solitary tract nucleus. Ascending endorphinergic pathways from the hypothalamus go to the preoptic field, septal nuclei, and amygdala, and descending pathways go to the periaqueductal gray matter, the blue nucleus, and the reticular formation. Endorphinergic neurons are involved in the central regulation of analgesia, they stimulate the release of growth hormone, prolactin and vasopressin.
Enkephalinergic neurons
Enkephalin is a 5 amino acid peptide that functions as an endogenous opiate receptor ligand. Enkephalinergic neurons are located in the superficial layer of the posterior horn of the spinal cord and the nucleus of the spinal tract of the trigeminal nerve, the perioval nucleus (auditory system), olfactory bulbs, in the nuclei of the raphe, in the gray periaqueductal substance. Enkephalin-containing neurons are also found in the neocortex and allocortex.
Enkephalinergic neurons presynaptically inhibit the release of substance P from the synaptic endings of afferents that conduct pain impulses (Fig. 1.39). Analgesia can be achieved by electrical stimulation or microinjection of opiates into this area. Enkephalinergic neurons affect the hypothalamic-pituitary regulation of the synthesis and release of oxytocin, vasopressin, some liberins and statins.
Nitric oxide
Nitric oxide (NO) is a multifunctional physiological regulator with the properties of a neurotransmitter, which, unlike traditional neurotransmitters, is not reserved in synaptic vesicles of nerve endings and is released into the synaptic cleft by free diffusion, and not by the mechanism of exocytosis. The NO molecule is synthesized in response to physiological need by the enzyme WA synthase (WAS) from the amino acid L-arginine. The ability of NO to produce a biological effect is determined mainly by the small size of its molecule, its high reactivity, and the ability to diffuse in tissues, including the nervous one. This was the basis for calling NO a retrograde messenger.
There are three forms of WAV. Two of them are constitutive: neuronal (ncNOS) and endothelial (ecWAS), the third is inducible (WAV), found in glial cells.
Calcium-calmodulin dependence of the neuronal WAV isoform causes an increase in NO synthesis with an increase in the level of intracellular calcium. In this regard, any processes leading to the accumulation of calcium in the cell (energy deficit, changes in active ion transport,
 Rice. 1.39. The mechanism of enkephalinergic regulation of pain sensitivity at the level of the gelatinous substance.
Rice. 1.39. The mechanism of enkephalinergic regulation of pain sensitivity at the level of the gelatinous substance.
1 - interneuron; 2 - enkephalin; 3 - enkephalin receptors; 4 - neuron of the posterior horn of the spinal cord; 5 - substance P receptors; 6 - substance P; 7 - sensitive neuron of the spinal ganglion. In the synapse between a peripheral sensory neuron and a neuron of the spinothalamic ganglion, substance P is the main mediator. The enkephalinergic interneuron responds to pain sensitivity, exerting a presynaptic inhibitory effect on the release of substance P
glutamate excitotoxicity, oxidative stress, inflammation) are accompanied by an increase in NO levels.
NO has been shown to have a modulating effect on synaptic transmission and on the functional state of NMDA glutamate receptors. By activating soluble heme-containing guanylate cyclase, NO is involved in the regulation of the intracellular concentration of Ca 2+ ions and pH inside nerve cells.
1.8. axonal transport
Axonal transport plays an important role in interneuronal connections. Membrane and cytoplasmic components, which are formed in the biosynthetic apparatus of the soma and the proximal part of the dendrites, must be distributed along the axon (their entry into the presynaptic structures of synapses is especially important) in order to compensate for the loss of elements that have undergone release or inactivation.
However, many axons are too long for materials to efficiently move from the soma to the synaptic terminals by simple diffusion. This task is performed by a special mechanism - axonal transport. There are several types of it. Membrane-surrounded organelles and mitochondria are transported at a relatively high rate via fast axonal transport. Substances dissolved in the cytoplasm (for example, proteins) move with the help of slow axonal transport. In mammals, fast axonal transport has a speed of 400 mm/day, and slow transport is about 1 mm/day. Synaptic vesicles can be transported by fast axonal transport from the motor neuron soma of the human spinal cord to the muscles of the foot in 2.5 days. Compare: the delivery of many soluble proteins over the same distance takes about 3 years.
Axonal transport requires the expenditure of metabolic energy and the presence of intracellular calcium. Elements of the cytoskeleton (more precisely, microtubules) create a system of guide strands along which organelles surrounded by membranes move. These organelles attach to microtubules in a manner similar to that between the thick and thin filaments of skeletal muscle fibers; the movement of organelles along microtubules is triggered by Ca 2+ ions.
Axonal transport occurs in two directions. Transport from the soma to axonal terminals, called anterograde axonal transport, replenishes the supply of synaptic vesicles and enzymes responsible for neurotransmitter synthesis in presynaptic endings. Transport in the opposite direction, retrograde axon transport, returns emptied synaptic vesicles to the soma, where these membrane structures are degraded by lysosomes. Substances coming from the synapses are necessary to maintain the normal metabolism of the bodies of nerve cells and, in addition, carry information about the state of their terminal apparatuses. Violation of retrograde axonal transport leads to changes in the normal functioning of nerve cells, and in severe cases, to retrograde degeneration of neurons.
The axonal transport system is the main mechanism that determines the renewal and supply of mediators and modulators in presynaptic endings, and also underlies the formation of new processes, axons and dendrites. According to the concept of brain plasticity in general, even in the adult brain, two interrelated processes constantly occur: the formation of new processes and synapses, as well as the destruction and disappearance of some part of the previously existing interneuronal contacts. The mechanisms of axonal transport, the processes of synaptogenesis associated with them, and the growth of the finest axon ramifications underlie learning, adaptation, and compensation for impaired functions. Disorder of axonal transport leads to the destruction of synaptic endings and changes in the functioning of certain brain systems.
medicinal and biological active substances it is possible to influence the metabolism of neurons, which determines their axonal transport, stimulating it and thereby increasing the possibility of compensatory-restorative processes. Strengthening axonal transport, the growth of the thinnest branches of axons and synaptogenesis play a positive role in the normal functioning of the brain. In pathology, these phenomena underlie reparative, compensatory and recovery processes.
Some viruses and toxins spread through axonal transport along the peripheral nerves. Yes, the varicella-zoster virus (Varicella zoster virus) penetrates into the cells of the spinal (spinal) ganglia. There, the virus remains in an inactive form, sometimes for many years, until the person's immune status changes. Then the virus can be transported along the sensory axons to the skin, and in the dermatomes
flaccid spinal nerves cause painful rashes of herpes zoster (Herpes zoster). Tetanus toxin is also transported by axonal transport. bacteria Clostridium tetani from a contaminated wound by retrograde transport enter the motor neurons. If the toxin is released into the extracellular space of the anterior horns of the spinal cord, it blocks the activity of synaptic inhibitory neurotransmitter amino acid receptors and causes tetanic convulsions.
1.9. Nervous tissue responses to injury
Damage to the nervous tissue is accompanied by reactions of neurons and neuroglia. When severely damaged, the cells die. Since neurons are post-mitotic cells, they do not replenish.
Mechanisms of death of neurons and glial cells
In severely damaged tissues, necrosis processes predominate, affecting entire cell fields with passive cell degeneration, swelling and fragmentation of organelles, membrane destruction, cell lysis, release of intracellular contents into the surrounding tissue, and development of an inflammatory response. Necrosis is always caused by a gross pathology, its mechanisms do not require energy expenditure, and it can only be prevented by removing the cause of the damage.
apoptosis is a type of programmed cell death. Apoptotic cells, in contrast to necrotic ones, are located singly or in small groups, scattered throughout the tissue. They have a smaller size, unchanged membranes, wrinkled cytoplasm with the preservation of organelles, the appearance of multiple cytoplasmic membrane-bound protrusions. There is no inflammatory reaction of the tissue, which is currently one of the important distinguishing morphological features of apoptosis from necrosis. Both shrunken cells and apoptotic bodies contain intact cell organelles and masses of condensed chromatin. The result of sequential DNA destruction in apoptotic cells is the impossibility of their replication (reproduction) and participation in intercellular interactions, since these processes require the synthesis of new proteins. Dying cells are effectively removed from the tissue by phagocytosis. The main differences between the processes of necrosis and apoptosis are summarized in Table 1. 1.1.
Table 1.1. Signs of differences in the processes of necrosis and apoptosis
 Apoptosis is an integral part of the processes of development and homeostasis of mature tissue. Normally, the body uses this genetically programmed mechanism during embryogenesis to destroy the “excess” of cellular material at an early stage of tissue development, in particular, in neurons that have not established contacts with target cells and are thus deprived of trophic support from these cells. In adulthood, the intensity of apoptosis in the CNS of mammals significantly decreases, although it remains high in other tissues. The elimination of virus-affected cells and the development of an immune response are also accompanied by an apoptotic reaction. Along with apoptosis, other variants of programmed cell death are also isolated.
Apoptosis is an integral part of the processes of development and homeostasis of mature tissue. Normally, the body uses this genetically programmed mechanism during embryogenesis to destroy the “excess” of cellular material at an early stage of tissue development, in particular, in neurons that have not established contacts with target cells and are thus deprived of trophic support from these cells. In adulthood, the intensity of apoptosis in the CNS of mammals significantly decreases, although it remains high in other tissues. The elimination of virus-affected cells and the development of an immune response are also accompanied by an apoptotic reaction. Along with apoptosis, other variants of programmed cell death are also isolated.
Morphological markers of apoptosis are apoptotic bodies and shrunken neurons with an intact membrane. A biochemical marker that has become almost identical to the concept of "apoptosis" is DNA fragmentation. This process is activated by Ca 2+ and Mg 2+ ions and inhibited by Zn 2+ ions. DNA cleavage occurs as a result of the action of calcium-magnesium-dependent endonuclease. It has been established that endonucleases cleave DNA between histone proteins, releasing fragments of regular length. The DNA is initially divided into large fragments of 50 and 300,000 bases, which are then cleaved into fragments of 180 base pairs, forming a "ladder" when separated by gel electrophoresis. DNA fragmentation does not always correlate with the morphology characteristic of apoptosis and is a conditional marker that is not equivalent to morphological criteria. The most perfect way to confirm apoptosis is the biological-histochemical method, which makes it possible to fix not only DNA fragmentation, but also an important morphological feature - apoptotic bodies.
The apoptosis program consists of three consecutive stages: making a decision about death or survival; implementation of the destruction mechanism; elimination of dead cells (degradation of cellular components and their phagocytosis).
The survival or death of cells is largely determined by the expression products of cW-family genes. The protein products of two of these genes, ced-3 And ced-4(“killer genes”) are essential for apoptosis to occur. Protein product of a gene ced-9 protects cells by preventing apoptosis by preventing gene firing ced-3 And ced-4. Other genes of the family ced encode proteins involved in the packaging and phagocytosis of dying cells, degradation of the DNA of a dead cell.
In mammals, homologues of the killer gene ced-3(and its protein products) are genes encoding interleukin-converting enzymes - caspases (cysteine aspartyl proteases), which have different substrate and inhibitory specificities. Inactive caspase precursors, procaspases, are present in all cells. Activation of procaspases in mammals is carried out by an analogue of the ced-4 gene - an excitatory factor of apoptotic protease-1 (Apaf-a), binding for ATP, which emphasizes the importance of the level of energy supply for the choice of the mechanism of death. When excited, caspases modify the activity of cellular proteins (polymerases, endonucleases, nuclear membrane components) responsible for DNA fragmentation in apoptotic cells. Activated enzymes begin DNA cleavage with the appearance of triphosphonucleotides at the breaks, causing the destruction of cytoplasmic proteins. The cell loses water and decreases, the pH of the cytoplasm decreases. The cell membrane loses its properties, the cell shrinks, and apoptotic bodies are formed. The process of rearrangement of cell membranes is based on the activation of syringomyelase, which cleaves the syringomyelin of the cell with the release of ceramide, which activates phospholipase A2. There is an accumulation of products of arachidonic acid. The proteins phosphatidylserine and vitronectin expressed during apoptosis are brought to the outer surface of the cell and signaled to macrophages that carry out phagocytosis of apoptotic bodies.
Nematode gene homologues ced-9, determining cell survival, in mammals is a family of proto-oncogenes bcl-2. AND bcl-2, and related protein bcl-x-l are present in the mammalian brain, where they protect neurons from apoptosis during ischemic exposure, removal of growth factors, and the influence of neurotoxins in vivo And in vitro. Analysis of bcl-2 gene expression products revealed a whole family of bcl-2-related proteins, including both anti-apoptotic (Bcl-2 And Bcl-x-l), and proapoptotic (Bcl-x-s, Bax, Bad, Bag) proteins. The proteins bax and bad have a homologous sequence and form heterodimers with bcl-2 And bcl-xl in vitro. For activity that suppresses death, bcl-2 And bcl-x-l must form dimers with protein bah, and dimers with the bad protein increase death. This led to the conclusion that bcl-2 and related molecules are key determinants of cell survival or cell death in the CNS. Molecular genetic studies have shown that
called gene family bcl-2, consisting of 16 genes with opposite functions, in humans it is mapped on chromosome 18. Anti-apoptotic effects are produced by six genes of the family, similar to the progenitor of the group bcl-2; the other 10 genes support apoptosis.
Pro- and anti-apoptotic effects of activated gene expression products bcl-2 realized through the modulation of mitochondrial activity. Mitochondria is a key player in apoptosis. It contains cytochrome C, ATP, Ca 2+ ions and apoptosis-inducing factor (AIF) - components necessary for the induction of apoptosis. The release of these factors from the mitochondria occurs when its membrane interacts with activated proteins of the family bcl-2, which are attached to the outer mitochondrial membrane at the points of convergence of the outer and inner membranes - in the region of the so-called permeabilization pore, which is a megachannel up to 2 nm in diameter. When attaching proteins bcl-2 to the outer membrane of the mitochondria, the megachannels of the pore expand to 2.4–3 nm. Through these channels, cytochrome C, ATP and AIF enter the cytosol of the cell from the mitochondria. Anti-apoptotic proteins of the family bcl-2, on the contrary, they close the megachannels, interrupting the progression of the apoptotic signal and protecting the cell from apoptosis. During apoptosis, mitochondria do not lose their integrity and are not destroyed. Released from the mitochondria, cytochrome C forms a complex with the apoptotic protease activating factor (APAF-l), caspase-9, and ATP. This complex is an apoptosome in which caspase-9 is activated, and then the main "killer" caspase-3, which leads to cell death. The mitochondrial signaling mechanism is the main pathway for apoptosis induction.
Another mechanism of apoptosis induction is the transmission of a proapoptotic signal when the ligand binds to the receptors of the cell death region, which occurs through the adapter proteins FADD/MORT1, TRADD. The receptor pathway of cell death is much shorter than the mitochondrial one: by means of adapter molecules, caspase-8 is activated, which, in turn, directly activates "killer" caspases.
Certain proteins such as p53, p21 (WAF1), may promote apoptosis. It has been shown that natural p53 induces apoptosis in tumor cell lines and in vivo. Transformation p53 from the natural type to the mutant form leads to the development of cancer in many organs as a result of the suppression of apoptosis processes.
Axon degeneration
After transection of the axon in the soma of the nerve cell, the so-called axon reaction develops, aimed at restoring the axon by synthesizing new structural proteins. In the soma of intact neurons, Nissl bodies are intensely stained with the basic aniline dye, which binds to the ribonucleic acids of the ribosomes. However, during the axon reaction, the cisterns of the rough endoplasmic reticulum increase in volume, being filled with protein synthesis products. Chromatolysis occurs - the disorganization of ribosomes, as a result of which the staining of Nissl bodies with the main aniline dye becomes much weaker. The cell body swells and rounds, and the nucleus shifts to one side (eccentric position of the nucleus). All these morphological changes are a reflection of the cytological processes accompanying increased protein synthesis.
The section of the axon distal to the site of transection dies. Within a few days, this site and all synaptic endings of the axon are destroyed. The myelin sheath of the axon also degenerates, its fragments are captured by phagocytes. However, neuroglial cells that form myelin do not die. This sequence of phenomena is called Wallerian degeneration.
If the damaged axon provided the only or main synaptic input to the nerve or effector cell, then the postsynaptic cell may degenerate and die. A well-known example is the atrophy of skeletal muscle fibers after a violation of their innervation by motor neurons.
Axon regeneration
After the damaged axon degenerates, many neurons can grow a new axon. At the end of the proximal segment, the axon begins to branch [sprouting (sprouting)- growth]. In the PNS, newly formed branches grow along the original path of the dead nerve, if, of course, this path is available. During the period of Wallerian degeneration, the Schwann cells of the distal part of the nerve not only survive, but also proliferate, lining up in rows where the dead nerve passed. The "growth cones" of the regenerating axon make their way between the rows of Schwann cells and eventually can reach their targets, reinnervating them. The axons are then remyelinated by Schwann cells. The regeneration rate is limited
is measured by the rate of slow axon transport, i.e. about 1 mm/day.
Axonal regeneration in the CNS is somewhat different: oligodendroglia cells cannot chart a path for axonal branch growth because in the CNS each oligodendrocyte myelinates many axons (unlike the Schwann cells in the PNS, each of which supplies only one axon with myelin).
It is important to note that chemical signals have different effects on regenerative processes in the CNS and PNS. An additional obstacle to axonal regeneration in the CNS is glial scars formed by astrocytes.
Synaptic sprouting, which ensures the "re-enhancement" of existing neuronal currents and the formation of new polysynaptic connections, determines the plasticity of neuronal tissue and forms the mechanisms involved in the restoration of impaired neurological functions.
Trophic factors
An important role in the development of ischemic damage to brain tissue is played by the level of its trophic supply.
Neurotrophic properties are inherent in many proteins, including structural proteins (for example, S1OOβ). At the same time, they are maximized by growth factors, which represent a heterogeneous group of trophic factors, consisting of at least 7 families - neurotrophins, cytokines, fibroblast growth factors, insulin-dependent growth factors, the family of transforming growth factor 31 (TGF-J3I), epidermal growth factors and others, including growth protein 6 (GAP-6)4, platelet-dependent growth factor, heparin-bound neurotrophic factor, erythropoietin, macrophage colony-stimulating factor, etc. (Table 1.2).
The strongest trophic influence on all the main processes of the vital activity of neurons is exerted by neurotrophins - regulatory proteins of the nervous tissue, which are synthesized in its cells (neurons and glia). They act locally - at the site of release and especially intensively induce branching of dendrites and growth of axons in the direction of target cells.
To date, three neurotrophins that are similar in structure to each other have been most studied: nerve growth factor (NGF), brain-derived growth factor (BDNF), and neurotrophin-3 (NT-3).
Table 1.2. Modern classification of neurotrophic factors

 In a developing organism, they are synthesized by a target cell (for example, a muscle spindle), diffuse towards a neuron, and bind to receptor molecules on its surface.
In a developing organism, they are synthesized by a target cell (for example, a muscle spindle), diffuse towards a neuron, and bind to receptor molecules on its surface.
Receptor-bound growth factors are taken up by neurons (i.e. undergo endocytosis) and transported retrogradely into the soma. There they can act directly on the nucleus, changing the formation of enzymes responsible for the synthesis of neurotransmitters and the growth of axons. There are two forms of receptors for growth factors - low-affinity receptors and high-affinity tyrosine kinase receptors, to which most trophic factors bind.
As a result, the axon reaches the target cell, establishing synaptic contact with it. Growth factors support the life of neurons, which in their absence cannot exist.
Trophic dysregulation is one of the universal components of the pathogenesis of damage to the nervous system. When mature cells are deprived of trophic support, biochemical and functional dedifferentiation of neurons develops with a change in the properties of innervated tissues. Trophic dysregulation affects the state of macromolecules involved in membrane electrogenesis, active ion transport, synaptic transmission (enzymes for the synthesis of mediators, postsynaptic receptors) and effector function (muscle myosin). Ensembles of dedifferentiated central neurons create foci of pathologically enhanced excitation, triggering pathobiochemical cascades that lead to neuron death through the mechanisms of necrosis and apoptosis. On the contrary, with a sufficient level of trophic supply, regression of the neurological deficit after ischemic brain damage is often observed even with the remaining morphological defect that initially caused it, which indicates a high adaptability of the brain function.
It has been established that changes in potassium and calcium homeostasis, excessive synthesis of nitric oxide, which blocks the tyrosine kinase enzyme, which is part of the active center of trophic factors, and imbalance of cytokines, take part in the development of trophic deficiency. One of the proposed mechanisms is autoimmune aggression against its own neurotrophins and structural neurospecific proteins with trophic properties, which becomes possible as a result of a violation of the protective function of the blood-brain barrier.
The cerebellum is central authority balance and coordination of movements. It is formed by two hemispheres with a large number of grooves and convolutions, and a narrow middle part - a worm.
The bulk of the gray matter in the cerebellum is located on the surface and forms its cortex. A smaller part of the gray matter lies deep in the white matter in the form of the central nuclei of the cerebellum.
There are 3 layers in the cerebellar cortex: 1) the outer molecular layer contains relatively few cells, but many fibers. It distinguishes between basket and stellate neurons, which are inhibitory. Star-shaped - slow down vertically, basket-shaped - send axons over long distances, which end on the bodies of pear-shaped cells. 2) The middle ganglionic layer is formed by one row of large pear-shaped cells, first described by the Czech scientist Jan Purkinje. The cells have a large body, 2-3 short dendrites extend from the top, which branch in a small layer. 1 axon departs from the base, which goes into the white matter to the cerebellar nuclei. 3) The inner granular layer is characterized by a large number of tightly lying cells. Among the neurons, granule cells, Golgi cells (stellate), and fusiform horizontal neurons are distinguished here. Granule cells are small cells that have short dendrites, the latter forming excitatory synapses with mossy fibers in the cerebellar glamelurs. The granule cells excite the mossy fibers, and the axons go into the molecular layer and transmit information to the piriform cells and all the fibers located there. It is the only excitatory neuron in the cerebellar cortex. Golgi cells lie under the bodies of pear-shaped neurons, axons go to the cerebellar glameruli, and can inhibit impulses from mossy fibers to granule cells.
Afferent pathways enter the cerebellar cortex through 2 types of fibers: 1) liana-shaped (climbing) - they rise from the white matter through the granular and ganglionic layers. They reach the molecular layer, form synapses with the dendrites of pear-shaped cells and excite them. 2) Bryophytes - from the white matter they enter the granular layer. Here they form synapses with the dendrites of granular cells, and the axons of granular cells go into the molecular layer, forming synapses with the dendrites of pear-shaped neurons, which form inhibitory nuclei.
The cerebral cortex. Development, neural composition and layered organization. The concept of cyto- and myeloarchitectonics. Blood-brain barrier. Structural and functional unit of the cortex.
The cerebral cortex is the highest and most complexly organized nerve center of the screen type, whose activity ensures the regulation of various body functions and complex forms of behavior. The cortex is made up of a layer of gray matter. Gray matter contains nerve cells, nerve fibers, and neuroglial cells.
Among the multipolar neurons of the cortex, pyramidal, stellate, fusiform, arachnid, horizontal, "candelabra" cells, cells with a double bouquet of dendrites, and some other types of neurons are distinguished.
Pyramidal neurons constitute the main and most specific form for the cortex of the hemispheres. They have an elongated cone-shaped body, the apex of which faces the surface of the cortex. Dendrites extend from the apex and lateral surfaces of the body. Axons originate from the base of the pyramidal cells.
Pyramidal cells of different layers of the cortex differ in size and have different functional significance. Small cells are intercalary neurons. The axons of the large pyramids take part in the formation of motor pyramidal pathways.
The neurons of the cortex are located in unsharply demarcated layers, which are designated by Roman numerals and numbered from outside to inside. Each layer is characterized by the predominance of any one type of cell. There are six main layers in the cerebral cortex:
I - The molecular layer of the cortex contains a small number of small associative horizontal Cajal cells. Their axons run parallel to the surface of the brain as part of the tangential plexus of nerve fibers of the molecular layer. However, the bulk of the fibers of this plexus is represented by branching of the dendrites of the underlying layers.
II - The outer granular layer is formed by numerous small pyramidal and stellate neurons. The dendrites of these cells rise into the molecular layer, and the axons either go into the white matter, or, forming arcs, also enter the tangential plexus of fibers of the molecular layer.
III - The widest layer of the cerebral cortex is pyramidal. It contains pyramidal neurons, and spindle cells. The apical dendrites of the pyramids go into the molecular layer, the lateral dendrites form synapses with adjacent cells of this layer. The axon of the pyramidal cell always departs from its base. In small cells, it remains within the cortex; in large cells, it forms a myelin fiber that goes to the white matter of the brain. Axons of small polygonal cells are sent to the molecular layer. The pyramidal layer performs mainly associative functions.
IV - The inner granular layer in some areas of the cortex is very strongly developed (for example, in the visual and auditory cortex), while in others it may be almost absent (for example, in the precentral gyrus). This layer is formed by small stellate neurons. It consists of a large number of horizontal fibers.
V - The ganglionic layer of the cortex is formed by large pyramids, and the region of the motor cortex (precentral gyrus) contains giant pyramids, which were first described by the Kyiv anatomist V. A. Bets. The apical dendrites of the pyramids reach the 1st layer. The axons of the pyramids are projected to the motor nuclei of the brain and spinal cord. The longest axons of Betz cells in the pyramidal pathways reach the caudal segments of the spinal cord.
VI - The layer of polymorphic cells is formed by neurons of various shapes (fusiform, stellate). The axons of these cells go into the white matter as part of the efferent pathways, and the dendrites reach the molecular layer.
Cytoarchitectonics - features of the location of neurons in different parts of the cerebral cortex.
Among the nerve fibers of the cerebral cortex, one can single out associative fibers that connect individual parts of the cortex of one hemisphere, commissural fibers that connect the cortex of different hemispheres, and projection fibers, both afferent and efferent, that connect the cortex with the nuclei of the lower parts of the central nervous system.
autonomic nervous system. General structural characteristics and main functions. The structure of sympathetic and parasympathetic reflex arcs. Differences between vegetative reflex arcs and somatic ones.
The nervous system is divided into somatic, which innervates the skeletal muscle tissue (meaningful motor processes) and the autonomic nervous system, which regulates the function of internal organs, glands and blood vessels (unconscious regulation). It distinguishes the sympathetic and parasympathetic systems that regulate visceral functions.
Thus, the nervous system regulates and coordinates the functions of organs and systems as a whole.
Sympathetic nuclei are located in the spinal cord. The nerve fibers departing from it end outside the spinal cord in the sympathetic ganglions, from which the nerve fibers originate. These fibers are suitable for all organs.
Parasympathetic nuclei lie in the middle and medulla oblongata and in the sacral part of the spinal cord. Nerve fibers from the nuclei of the medulla oblongata are part of the vagus nerves. From the nuclei of the sacral part, the nerve fibers go to the intestines, the excretory organs.
The sympathetic nervous system enhances metabolism, increases the excitability of most tissues, and mobilizes the body's forces for vigorous activity. The parasympathetic system contributes to the restoration of spent energy reserves, regulates the functioning of the body during sleep.
The autonomic and somatic nervous systems, as a rule, act in concert, since any motor reaction requires autonomic accompaniment, which ensures the proper level of blood supply to the working muscles. Morphologically, it is impossible to distinguish between the nerve centers of the autonomic and somatic systems, but their peripheral parts are completely different.
The main morphological difference between the autonomic nervous system and the somatic one is the two-neuronality of the efferent (centrifugal) pathway: the processes of the central autonomic neurons themselves do not reach the structures of the innervated organ, but switch to the second neuron, the endings of which already directly innervate the controlled structures.
Similar information.
To control the work of internal organs, motor functions, timely receipt and transmission of sympathetic and reflex impulses, the pathways of the spinal cord are used. Violations in the transmission of impulses leads to serious malfunctions in the work of the whole organism.
What is the conduction function of the spinal cord
The term "conducting pathways" means a set of nerve fibers that provide signal transmission to various centers of gray matter. The ascending and descending tracts of the spinal cord perform the main function - the transmission of impulses. It is customary to distinguish three groups of nerve fibers:- Associative pathways.
- Commissary connections.
- Projective nerve fibers.
Sensory and motor pathways provide a strong relationship between the spinal cord and brain, internal organs, muscular system and musculoskeletal system. Due to the rapid transmission of impulses, all body movements are carried out in a coordinated manner, without tangible effort on the part of the person.
What are the conducting tracts of the spinal cord formed by?
The main pathways are formed by bundles of cells - neurons. This structure provides the necessary speed of pulse transmission.The classification of the pathways depends on the functional characteristics of the nerve fibers:
- Ascending pathways of the spinal cord - read and transmit signals: from the skin and mucous membranes of a person, life-support organs. Ensure the performance of the functions of the musculoskeletal system.
- Descending pathways of the spinal cord - transmit impulses directly to the working organs of the human body - muscle tissues, glands, etc. Connected directly to the cortical part of the gray matter. The transmission of impulses occurs through the spinal neural connection to the internal organs.
The spinal cord has a double direction of conducting paths, which provides a fast impulse transmission of information from controlled organs. The conductive function of the spinal cord is carried out due to the presence of an effective transmission of impulses through the nervous tissue.
In medical and anatomical practice, it is customary to use the following terms: 
Where are the pathways of the spinal cord located?
All nervous tissues are located in the gray and white matter, connect the spinal horns and the cerebral cortex.The morphofunctional characteristic of the descending pathways of the spinal cord limits the direction of impulses in only one direction. Synapses are irritated from the presynaptic to the postsynaptic membrane.
The conduction function of the spinal cord and brain corresponds to the following possibilities and the location of the main ascending and descending pathways:
- Associative pathways - are "bridges" connecting the areas between the cortex and the nuclei of gray matter. Composed of short and long fibers. The first are located within one half or lobe of the cerebral hemispheres.
Long fibers are able to transmit signals through 2-3 segments of the gray matter. In the cerebrospinal substance, neurons form intersegmental bundles. - Commissural fibers - form the corpus callosum, connecting the newly formed sections of the spinal cord and brain. Disperse in a radiant way. They are located in the white matter of the brain tissue.
- Projection fibers - the location of the pathways in the spinal cord allows impulses to reach the cerebral cortex as quickly as possible. By their nature and functional features, the projection fibers are divided into ascending (afferent pathways) and descending.
The first are divided into exteroceptive (vision, hearing), proprioceptive (motor functions), interoreceptive (communication with internal organs). The receptors are located between the spinal column and the hypothalamus.

The anatomy of the pathways is quite complicated for a person who does not have medical education. But neural transmission of impulses is what makes the human body a single whole.
The consequences of damage to the pathways
To understand the neurophysiology of the sensory and motor pathways, it is necessary to become familiar with the anatomy of the spine. The spinal cord has a structure much like a cylinder surrounded by muscle tissue. Inside the gray matter are conductive paths that control the functioning of internal organs, as well as motor functions. Associative pathways are responsible for pain and tactile sensations. Motor - for the reflex functions of the body.
Inside the gray matter are conductive paths that control the functioning of internal organs, as well as motor functions. Associative pathways are responsible for pain and tactile sensations. Motor - for the reflex functions of the body.
As a result of trauma, malformations or diseases of the spinal cord, conduction may decrease or stop completely. This happens due to the death of nerve fibers. For a complete violation of the conduction of impulses of the spinal cord is characterized by paralysis, lack of sensitivity of the limbs. Failures in the work of internal organs begin, for which the damaged neural connection is responsible. So, with damage to the lower part of the spinal cord, urinary incontinence and spontaneous defecation are observed.
The reflex and conduction activity of the spinal cord is disturbed immediately after the onset of degenerative pathological changes. There is a death of nerve fibers that are difficult to restore. The disease progresses rapidly and a gross violation of conduction occurs. For this reason, it is necessary to start medical treatment as early as possible.
How to restore patency in the spinal cord
The treatment of non-conductivity is primarily associated with the need to stop the death of nerve fibers, as well as to eliminate the causes that have become a catalyst for pathological changes.Medical treatment
It consists in the appointment of drugs that prevent the death of brain cells, as well as sufficient blood supply to the damaged area of the spinal cord. This takes into account the age-related features of the conductive function of the spinal cord, as well as the severity of the injury or disease.For additional stimulation of nerve cells, electrical impulse treatment is performed to help maintain muscle tone.
Surgery
The operation to restore the conduction of the spinal cord affects two main areas:- Elimination of catalysts that caused the paralysis of neural connections.
- Spinal cord stimulation to restore lost functions.
Traditional medicine for conduction disorders
Folk remedies for impaired conduction of the spinal cord, if used, should be used with extreme caution so as not to worsen the patient's condition.Particularly popular are: 
It is quite difficult to fully restore neural connections after an injury. Much depends on a quick appeal to a medical center and qualified assistance from a neurosurgeon. The more time passes from the onset of degenerative changes, the less chance there is to restore the functionality of the spinal cord.