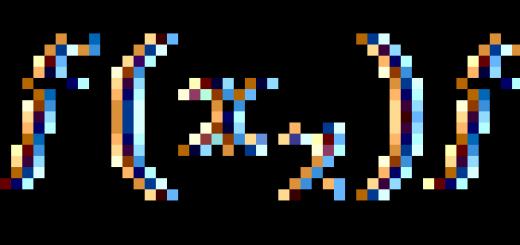Belgorod regional public organization
MBOUDOD "Center for children's and youth tourism and excursions"
G. Belgorod
Methodical development
Topic:"Physiological basis of adaptation of the athlete's body to new climatic conditions"
trainer-teacher TsDYUTE
Belgorod, 2014
1. Concept of adaptation
2. Adaptation and homeostasis
3. Cold adaptation
4. Acclimatization. mountain sickness
5. The development of specific endurance as a factor contributing to high-altitude acclimatization
1. Concept of adaptation
AdaptationIt is a process of adaptation that is formed during a person's life. Thanks to adaptive processes, a person adapts to unusual conditions or a new level of activity, i.e., the resistance of his body against the action of various factors increases. The human body can adapt to high and low temperatures, emotional stimuli (fear, pain, etc.), low atmospheric pressure, or even some pathogenic factors.
For example, a climber adapted to a lack of oxygen can climb a mountain peak with a height of 8000 m or more, where the partial pressure of oxygen approaches 50 mm Hg. Art. (6.7 kPa). The atmosphere at such an altitude is so rarefied that an untrained person dies in a few minutes (due to lack of oxygen) even at rest.
People living in the northern or southern latitudes, in the mountains or on the plains, in the humid tropics or in the desert differ from each other in many indicators of homeostasis. Therefore, a number of normal indicators for individual regions of the globe may differ.
We can say that human life in real conditions is a constant adaptation process. Its body adapts to the effects of various climatic and geographical, natural (atmospheric pressure and gas composition of air, duration and intensity of insolation, temperature and humidity, seasonal and daily rhythms, geographical longitude and latitude, mountains and plains, etc.) and social factors, conditions of civilization . As a rule, the body adapts to the action of a complex of various factors.The need to stimulate the mechanisms that drive the process of adaptation arises as the strength or duration of the impact of a number of external factors increases. For example, in the natural conditions of life, such processes develop in autumn and spring, when the body is gradually rebuilt, adapting to cold weather or warming.
Adaptation also develops when a person changes the level of activity and begins to engage in physical education or some uncharacteristic type of labor activity, i.e., the activity of the motor apparatus increases. In modern conditions, in connection with the development of high-speed transport, a person often changes not only climatic and geographical conditions, but also time zones. This leaves its mark on biorhythms, which is also accompanied by the development of adaptive processes.
2. Adaptation and homeostasis
A person is forced to constantly adapt to changing environmental conditions, preserving his body from destruction under the influence of external factors. The preservation of the body is possible due to homeostasis - a universal property to preserve and maintain the stability of the work of various body systems in response to influences that violate this stability.
homeostasis- relative dynamic constancy of the composition and properties of the internal environment and the stability of the basic physiological functions of the body. Any physiological, physical, chemical or emotional influences, be it air temperature, changes in atmospheric pressure or excitement, joy, sadness, can be a reason for the body to get out of a state of dynamic equilibrium. Automatically, with the help of humoral and nervous mechanisms of regulation, self-regulation of physiological functions is carried out, which ensures the maintenance of the vital activity of the organism at a constant level. Humoral regulation is carried out through the liquid internal environment of the body with the help of chemical molecules released by cells or certain tissues and organs (hormones, enzymes, etc.). Nervous regulation provides fast and directed transmission of signals in the form of nerve impulses arriving at the object of regulation.
Reactivity is an important property of a living organism that affects the efficiency of regulatory mechanisms. Reactivity is the ability of an organism to respond (react) with changes in metabolism and function to stimuli of the external and internal environment. Compensation for changes in environmental factors is possible due to the activation of systems responsible for adaptation(adaptation) of the organism to external conditions.
Homeostasis and adaptation are the two end results that organize functional systems. The intervention of external factors in the state of homeostasis leads to an adaptive restructuring of the body, as a result of which one or more functional systems compensate for possible disturbances and restore balance.
3. Cold adaptation
In the high mountains, under conditions of increased physical exertion, the most significant processes are acclimatization - adaptation to cold.
The optimal microclimatic zone corresponds to the temperature range of 15...21 °С; it ensures a person's well-being and does not cause shifts in thermoregulation systems;
The permissible microclimatic zone corresponds to the temperature range from minus 5.0 to plus 14.9°C and 21.7...27.0°C; ensures the preservation of human health for a long time of exposure, but causes discomfort, as well as functional shifts that do not go beyond the limits of its physiological adaptive capabilities. When in this zone, the human body is able to maintain a temperature balance due to changes in skin blood flow and sweating for a long time without deteriorating health;
Maximum permissible microclimatic zone, effective temperatures from 4.0 to minus 4.9°С and from 27.1 to 32.0°С. Maintaining a relatively normal functional state for 1-2 hours is achieved due to the tension of the cardiovascular system and the thermoregulation system. Normalization of the functional state occurs after 1.0-1.5 hours of stay in an optimal environment. Frequent repeated exposures lead to disruption of bulk processes, depletion of the body's defenses, and a decrease in its nonspecific resistance;
Extremely tolerable microclimatic zone, effective temperatures from minus 4.9 to minus 15.0 ºС and from 32.1 to 38.0 °С.
Performance of loading at temperatures in the specified ranges results in 30-60 min. to a pronounced change in the functional state: at low temperatures it is cool in fur clothes, hands in fur gloves freeze: at high temperatures, the heat sensation is “hot”, “very hot”, lethargy, unwillingness to work, headache, nausea, increased irritability appear; sweat, abundantly flowing from the forehead, gets into the eyes, interferes; with an increase in symptoms of overheating, vision is impaired.
The dangerous microclimatic zone below minus 15 and above 38 ° C is characterized by such conditions that after 10-30 minutes. May lead to poor health.
Uptime
when performing a load in adverse microclimatic conditions
Microclimate zone | Below optimal temperatures | Above optimal temperatures |
||
Effective temperature, С | Time, min. | Effective temperature, С | Time, min. |
|
Permissible | 5,0…14,9 | 60 – 120 | 21,7…27,0 | 30 – 60 |
Maximum allowable | From 4.9 to minus 4.9 | 30 – 60 | 27,1…32,0 | 20 – 30 |
Extremely portable | Minus 4.9…15.0 | 10 – 30 | 32,1…38,0 | 10 – 20 |
dangerous | Below minus 15.1 | 5 – 10 | Above 38.1 | 5 – 10 |
4 . Acclimatization. mountain sickness
As you go up in altitude, air pressure drops. Accordingly, the pressure of all components of air, including oxygen, drops. This means that the amount of oxygen entering the lungs during inhalation is less. And oxygen molecules are less intensively attached to blood erythrocytes. The concentration of oxygen in the blood decreases. The lack of oxygen in the blood is called hypoxia. Hypoxia leads to the development mountain sickness.
Typical manifestations of altitude sickness:
· increased heart rate;
· shortness of breath on exertion;
· headache, insomnia;
· weakness, nausea and vomiting;
· inappropriate behaviour.
In advanced cases, mountain sickness can lead to serious consequences.
To be safe at high altitudes, you need acclimatization- adaptation of the body to high altitude conditions.
Acclimatization is impossible without altitude sickness. Mild forms of mountain sickness trigger the body's restructuring mechanisms.
There are two phases of acclimatization:
· Short term acclimatization is a rapid response to hypoxia. The changes mainly concern oxygen transport systems. The frequency of respiration and heartbeat increases. Additional erythrocytes are ejected from the blood depot. There is a redistribution of blood in the body. Increases cerebral blood flow, because the brain requires oxygen. This is what leads to headaches. But such adaptation mechanisms can only be effective for a short time. At the same time, the body experiences stress and wears out.
· Long-term acclimatization - a complex of profound changes in the body. It is she who is the purpose of acclimatization. In this phase, the focus shifts from transport mechanisms to mechanisms for the economical use of oxygen. The capillary network grows, the area of the lungs increases. The composition of the blood changes - embryonic hemoglobin appears, which more easily attaches oxygen at its low partial pressure. The activity of enzymes that break down glucose and glycogen increases. The biochemistry of myocardial cells changes, which allows more efficient use of oxygen.
Step acclimatization
When climbing to a height, the body experiences a lack of oxygen. Mild mountain sickness sets in. Mechanisms of short-term acclimatization are included. For effective acclimatization after the ascent, it is better to go down, so that changes in the body occur in more favorable conditions and there is no exhaustion of the body. This is the principle of stepwise acclimatization - a sequence of ascents and descents, in which each subsequent ascent is higher than the previous one.
Rice. 1. Sawtooth graph of stepwise acclimatization
Sometimes the features of the relief do not provide an opportunity for a full-fledged stepwise acclimatization. For example, on many tracks in the Himalayas, where climbing takes place daily. Then daytime transitions are made small so that the height increase does not occur too quickly. It is very useful in this case to look for an opportunity to make even a small exit up from the place of spending the night. Often you can take a walk in the evening on a nearby hill or a spur of a mountain, and gain at least a couple of hundred meters.
What should be done to ensure successful acclimatization before the trip?
General physical training . It is easier for a trained athlete to endure the loads associated with height. First of all, you should develop endurance. This is achieved by sustained low-intensity exercise. The most accessible means of developing endurance is run.
It is practically useless to run often, but little by little. It is better to run once a week for 1 hour than every day for 10 minutes. For the development of endurance, the length of the runs should be more than 40 minutes, the frequency - according to the sensations. It is important to monitor the pulse rate and not overload the heart. In general, training should be enjoyable, fanaticism is not needed.
Health.It is very important to come to the mountains healthy and rested. If you have been training, then three weeks before the trip, reduce the load and give the body a rest. Adequate sleep and nutrition are required. Nutrition can be supplemented with vitamins and minerals. Minimize or better avoid alcohol. Avoid stress and overwork at work. You need to fix your teeth.
In the early days, the body is subject to heavy loads. The immune system weakens and it is easy to get sick. Avoid hypothermia or overheating. In the mountains, there are sharp temperature changes and therefore you need to follow the rule - undress before you sweat, dress before you get cold.
Appetite at altitude can be reduced, especially if you immediately go to high altitudes. There is no need to force. Give preference to easily digestible foods. In the mountains, due to the dryness of the air and heavy physical exertion, a person needs a large amount of water - drink a lot.
Continue taking vitamins and minerals. You can start taking amino acids that have adaptogenic properties.
Movement mode.It happens that only after arriving in the mountains, tourists, experiencing an emotional upsurge and feeling overwhelmed by their strength, go too fast along the path. You need to restrain yourself, the pace of movement should be calm and uniform. In the early days in the highlands, the pulse at rest is 1.5 times higher than in the plains. It’s already hard for the body, so you don’t need to drive, especially on the climbs. Small tears may not be noticeable, but tend to accumulate, and can lead to a breakdown in acclimatization.
If you come to the place of spending the night, and you do not feel well, you do not need to go to bed. It is better to walk at a calm pace around the neighborhood, take part in the arrangement of the bivouac, in general, do something.
Movement and work - an excellent cure for mild forms of mountain sickness. Night is a very important time for acclimatization. Sleep must be sound. If you have a headache in the evening, take a painkiller. Headache destabilizes the body and cannot be tolerated. If you can't sleep, take sleeping pills. You can't stand insomnia either.
Check your heart rate before bed and in the morning immediately after waking up. The morning pulse should be lower - this is an indicator that the body has rested.
With well-planned preparation and the right climb schedule, you can avoid serious manifestations of altitude sickness and enjoy the conquest of great heights.
5. Development of specific endurance as a factor contributing to high-altitude acclimatization
"If a climber (mountain tourist) in the off-season and pre-season increases his "oxygen ceiling" by swimming, running, cycling, skiing, rowing, he will ensure the improvement of his body, he will then be more successful in coping with great, but exciting difficulties when storming mountain peaks ".
This recommendation is both true and false. In the sense that it is, of course, necessary to prepare for the mountains. But cycling, rowing, swimming and other types of training give different “improvement of your body” and, accordingly, a different “oxygen ceiling”. When it comes to the motor acts of the body, one should clearly understand that there is no "movement in general" and any motor act is extremely specific. And from a certain level, the development of one physical quality always occurs at the expense of another: strength due to endurance and speed, endurance due to strength and speed.
When training for intensive work the consumption of oxygen and oxidation substrates in the muscles per unit time is so high that it is unrealistic to quickly replenish their reserves by increasing the work of transport systems. The sensitivity of the respiratory center to carbon dioxide is reduced, which protects the respiratory system from unnecessary overstrain.
Muscles capable of performing such a load actually work in autonomous mode, relying on their own resources. This does not eliminate the development of tissue hypoxia and leads to the accumulation of large amounts of underoxidized products. An important aspect of adaptive reactions in this case is the formation of tolerance, that is, resistance to pH shift. This is ensured by an increase in the capacity of the buffer systems of blood and tissues, an increase in the so-called. alkaline reserve of the blood. The power of the antioxidant system in the muscles also increases, which weakens or prevents lipid peroxidation of cell membranes - one of the main damaging effects of the stress response. The power of the anaerobic glycolysis system increases due to the increased synthesis of glycolytic enzymes, the reserves of glycogen and creatine phosphate, energy sources for ATP synthesis, increase.
When training for moderate work the growth of the vascular network in the muscles, heart, lungs, an increase in the number of mitochondria and a change in their characteristics, an increase in the synthesis of oxidative enzymes, an increase in erythropoiesis, leading to an increase in the oxygen capacity of the blood, can reduce the level of hypoxia or prevent it. With the systematic performance of moderate physical activity, accompanied by an increase in pulmonary ventilation, the respiratory center, on the contrary, increases sensitivity to CO 2 , which is due to a decrease in its content due to leaching from the blood during increased breathing.
Therefore, in the process of adaptation to intensive (as a rule, short-term) work, a different spectrum of adaptive adaptations develops in the muscles than to long-term moderate work. Therefore, for example, during hypoxia during diving, it becomes impossible to activate external respiration, which is typical for adaptation to high-altitude hypoxia or hypoxia during muscular work. And the struggle to maintain oxygen homeostasis is manifested in an increase in oxygen reserves carried under water. Consequently, the range of adaptive adaptations for different types of hypoxia is different, therefore, it is far from always useful for high mountains.
Table. The volume of circulating blood (BCC) and its components in athletes training endurance and untrained (L. Röcker, 1977). |
||
Indicators | Athletes | Not athletes |
BCC [l] | 6,4 | 5,5 |
BCC [ml/kg body weight] | 95,4 | 76,3 |
Volume of circulating plasma (CVV) [l] | 3,6 | 3,1 |
VCP [ml/kg body weight] | 55,2 | 43 |
Volume of circulating erythrocytes (VCE) [l] | 2,8 | 2,4 |
OCE [ml/kg body weight] | 40,4 | 33,6 |
Hematocrit [%] | 42,8 | 44,6 |
So, in untrained and in representatives of speed-strength sports, the total hemoglobin content in the blood is 10-12 g / kg (in women - 8-9 g / kg), and in endurance athletes - g / kg (in athletes - 12 g /kg).
Athletes who train endurance show increased utilization of lactic acid formed in the muscles. This is facilitated by an increased aerobic potential of all muscle fibers and a particularly high percentage of slow muscle fibers, as well as an increased mass of the heart. Slow muscle fibers, like the myocardium, are able to actively use lactic acid as an energy substrate. In addition, with the same aerobic loads (equal consumption of O 2 ) blood flow through the liver in athletes is higher than in untrained, which can also contribute to a more intensive extraction of lactic acid from the blood by the liver and its further conversion into glucose and glycogen. Thus, aerobic endurance training not only increases aerobic capacity, but also develops the ability to perform large long-term aerobic exercise without a significant increase in blood lactic acid.
It is obvious that in winter it is better to do skiing, in the off-season - long distance cross-country running. The lion's share of the physical preparation of those who are going to high mountains should be devoted to these trainings. Not so long ago, scientists broke spears about what kind of distribution of forces when running is optimal. Some believed that the variable, others - uniform. It really depends on the level of training.
Literature
1. Pavlov. - M., "Sails", 2000. - 282 p.
2. Human physiology in high altitude conditions: A guide to physiology. Ed. . - Moscow, Nauka, 1987, 520 p.
3. Somero J. Biochemical adaptation. M.: Mir, 19s
4. Oxygen transport system and endurance
5. A. Lebedev. Planning sports trips
- Specialty HAC RF03.00.16
- Number of pages 101
CHAPTER 1. MODERN CONCEPTS ON THE MECHANISM OF ADAPTATION OF THE ORGANISM TO COLD AND TOCOPHEROL DEFICIENCY.
1.1 New ideas about the biological functions of reactive oxygen species during adaptive transformations of metabolism.
1.2 Mechanisms of body adaptation to cold and the role of oxidative stress in this process.
1.3 Mechanisms of adaptation of the body to tocopherol deficiency and the role of oxidative stress in this process.
CHAPTER 2. MATERIAL AND RESEARCH METHODS.
2.1 Organization of the study.
2.1.1 Organization of experiments on the influence of cold.
2.1.2 Organization of experiments on the effect of tocopherol deficiency.
2.2 Research methods
2.2.1 Hematological parameters
2.2.2 Study of energy metabolism.
2.2.3 Study of oxidative metabolism.
2.3 Statistical processing of results.
CHAPTER 3. INVESTIGATION OF OXIDATIVE HOMEOSTASIS, BASIC MORPHOFUNCTIONAL PARAMETERS OF THE ORGANISM OF RATS AND ERYTHROCYTES UNDER LONG-TERM EXPOSURE TO COLD.
CHAPTER 4. INVESTIGATION OF OXIDATIVE HOMEOSTASIS, BASIC MORPHOFUNCTIONAL PARAMETERS OF THE ORGANISM OF RATS AND ERYTHROCYTES WITH LONG-TERM TOCOPHEROL DEFICIENCY.
Recommended list of dissertations
Physiological aspects of cellular and molecular patterns of adaptation of animal organisms to extreme situations 2013, Doctor of Biological Sciences Cherkesova, Dilara Ulubievna
Mechanisms of participation of tocopherol in adaptive transformations in the cold 2000, Doctor of Biological Sciences Kolosova, Natalia Gorislavovna
Features of the functioning of the hypothalamic-pituitary-reproductive system at the stages of ontogenesis and under the conditions of the use of geroprotectors 2010, Doctor of Biological Sciences Kozak, Mikhail Vladimirovich
Ecological and physiological aspects of the formation of adaptive mechanisms of mammals to hypothermia under experimental conditions 2005, candidate of biological sciences Solodovnikova, Olga Grigoryevna
Biochemical mechanisms of the antistress effect of α-tocopherol 1999, Doctor of Biological Sciences Saburova, Anna Mukhammadievna
Introduction to the dissertation (part of the abstract) on the topic "Experimental study of enzyme antioxidant systems during adaptation to prolonged exposure to cold and tocopherol deficiency"
Relevance of the topic. Recent studies have shown that the so-called reactive oxygen species, such as superoxide and hydroxyl radicals, hydrogen peroxide, and others, play an important role in the mechanisms of organism adaptation to environmental factors (Finkel, 1998; Kausalya and Nath, 1998). It has been established that these free-radical oxygen metabolites, which until recently were considered only as damaging agents, are signaling molecules and regulate adaptive transformations of the nervous system, arterial hemodynamics, and morphogenesis. (Luscher, Noll, Vanhoute, 1996; ; Groves, 1999; Wilder, 1998; Drexler, Homig, 1999). The main source of reactive oxygen species is a number of enzymatic systems of the epithelium and endothelium (NADP-oxidase, cyclooxygenase, lipoxygenase, xanthine oxidase), which are activated upon stimulation of chemo- and mechanoreceptors located on the luminal membrane of the cells of these tissues.
At the same time, it is known that with increased production and accumulation in the body of reactive oxygen species, that is, with the so-called oxidative stress, their physiological function can be transformed into a pathological one with the development of peroxidation of biopolymers and damage to cells and tissues as a result. (Kausalua & Nath 1998; Smith & Guilbelrt & Yui et al. 1999). Obviously, the possibility of such transformation is determined primarily by the rate of ROS inactivation by antioxidant systems. In this regard, of particular interest is the study of changes in reactive oxygen species inactivators - enzymatic antioxidant systems of the body, with prolonged exposure of the body to such extreme factors as cold and deficiency of the vitamin antioxidant - tocopherol, which are currently considered as endo- and exogenous inducers of oxidative stress.
Purpose and objectives of the study. The aim of the work was to study changes in the main enzymatic antioxidant systems during adaptation of rats to prolonged exposure to cold and tocopherol deficiency.
Research objectives:
1. To compare changes in indicators of oxidative homeostasis with changes in the main morphological and functional parameters of the body of rats and erythrocytes during prolonged exposure to cold.
2. To compare changes in indicators of oxidative homeostasis with changes in the main morphological and functional parameters of the body of rats and erythrocytes in tocopherol deficiency.
3. Carry out a comparative analysis of changes in oxidative metabolism and the nature of the adaptive reaction of the body of rats during prolonged exposure to cold and tocopherol deficiency.
Scientific novelty. It has been established for the first time that prolonged intermittent exposure to cold (+5°C for 8 hours a day for 6 months) causes a number of adaptive morphofunctional changes in the body of rats: acceleration of body weight gain, an increase in the content of spectrin and actin in erythrocyte membranes , increased activity of key enzymes of glycolysis, the concentration of ATP and ADP, as well as the activity of ATPases.
For the first time, it has been shown that oxidative stress plays an important role in the mechanism of development of adaptation to cold, a feature of which is an increase in the activity of the components of the antioxidant system - enzymes of the NADPH-generating pentose phosphate pathway of glucose breakdown, superoxide dismutase, catalase, and glutathione pyroxidase.
It has been shown for the first time that the development of pathological morphological and functional changes in tocopherol deficiency is associated with severe oxidative stress occurring against the background of reduced activity of the main antioxidant enzymes and enzymes of the pentose phosphate pathway of glucose breakdown.
It has been established for the first time that the result of metabolic transformations under the influence of environmental factors on the body depends on the adaptive increase in the activity of antioxidant enzymes and the associated severity of oxidative stress.
Scientific and practical significance of the work. The new facts obtained in the work expand the understanding of the mechanisms of adaptation of the body to environmental factors. The dependence of the result of adaptive transformations of metabolism on the degree of activation of the main enzymatic antioxidants was revealed, which indicates the need for directed development of the adaptive potential of this non-specific system of stress resistance of the body under changing environmental conditions.
The main provisions for defense:
1. Prolonged exposure to cold causes a complex of changes in the adaptive direction in the body of rats: an increase in resistance to the action of cold, which was expressed in the weakening of hypothermia; acceleration of body weight gain; increase in the content of spectrin and actin in erythrocyte membranes; an increase in the rate of glycolysis, an increase in the concentration of ATP and ADP; an increase in the activity of ATPases. The mechanism of these changes is associated with the development of oxidative stress in combination with an adaptive increase in the activity of the components of the antioxidant defense system - pentose-phosphate shunt enzymes, as well as the main intracellular antioxidant enzymes, primarily superoxide dismutase.
2. A long-term deficiency of tocopherol in the body of rats causes a persistent hypotrophic effect, damage to erythrocyte membranes, inhibition of glycolysis, a decrease in the concentration of ATP and ADP, and the activity of cellular ATPases. In the mechanism of development of these changes, insufficient activation of antioxidant systems - the NADPH-generating pentose-phosphate pathway and antioxidant enzymes, which creates conditions for the damaging action of reactive oxygen species, is essential.
Approbation of work. The results of the research were reported at a joint meeting of the Department of Biochemistry and the Department of Normal Physiology of the Altai State Medical Institute (Barnaul, 1998, 2000), at a scientific conference dedicated to the 40th anniversary of the Department of Pharmacology of the Altai State Medical University (Barnaul, 1997), at a scientific and practical conference" Modern Problems of Balneology and Therapy", dedicated to the 55th anniversary of the sanatorium "Barnaul" (Barnaul, 2000), at the II International Conference of Young Scientists of Russia (Moscow, 2001).
Similar theses in the specialty "Ecology", 03.00.16 VAK code
Study of the role of the glutathione system in the natural aging of erythrocytes produced under conditions of normal and intense erythropoiesis 2002, candidate of biological sciences Kudryashov, Alexander Mikhailovich
Indicators of the antioxidant system of erythrocytes in burn injury 1999, candidate of biological sciences Eremina, Tatyana Vladimirovna
Biochemical changes in mammalian membranes during hibernation and hypothermia 2005, Doctor of Biological Sciences Klichkhanov, Nisred Kadirovich
Study of the effect of thioctic acid on free radical homeostasis in rat tissues with pathologies associated with oxidative stress 2007, Candidate of Biological Sciences Anna Vitalievna Makeeva
Ratio between prooxidant and antioxidant systems in erythrocytes under immobilization stress in rats 2009, candidate of biological sciences Lapteva, Irina Azatovna
Dissertation conclusion on the topic "Ecology", Skuryatina, Yulia Vladimirovna
1. Long-term intermittent exposure to cold (+5°C for 8 hours a day for 6 months) causes a complex of adaptive changes in the body of rats: dissipation of the hypothermic reaction to cold, acceleration of body weight gain, an increase in the content of spectrin and actin in erythrocyte membranes, increased glycolysis, an increase in the total concentration of ATP and ADP and the activity of ATPases.
2. The state of adaptation of rats to prolonged intermittent exposure to cold corresponds to oxidative stress, which is characterized by increased activity of the components of enzymatic antioxidant systems - glucose-6-phosphate dehydrogenase, superoxide dismutase, catalase and glutathione peroxidase.
3. Long-term (6 months) alimentary deficiency of tocopherol causes a persistent hypotrophic effect in the body of rats, anemia, damage to erythrocyte membranes, inhibition of glycolysis in erythrocytes, a decrease in the total concentration of ATP and ADP, as well as the activity of Na+,K+-ATPase.
4. Disadaptive changes in the body of rats with tocopherol deficiency are associated with the development of pronounced oxidative stress, which is characterized by a decrease in the activity of catalase and glutathione peroxidase, combined with a moderate increase in the activity of glucose-6-phosphate dehydrogenase and superoxide dismutase.
5. The result of adaptive transformations of metabolism in response to prolonged exposure to cold and alimentary tocopherol deficiency depends on the severity of oxidative stress, which is largely determined by an increase in the activity of antioxidant enzymes.
CONCLUSION
To date, a fairly clear idea has developed that the adaptation of the human and animal organism is determined by the interaction of the genotype with external factors (Meyerson and Malyshev, 1981; Panin, 1983; Goldstein and Brown, 1993; Ado and Bochkov, 1994). At the same time, it should be taken into account that the genetically determined inadequacy of the inclusion of adaptive mechanisms under the influence of extreme factors can lead to the transformation of a state of stress into an acute or chronic pathological process (Kaznacheev, 1980).
The process of adaptation of the organism to new conditions of the internal and external environment is based on the mechanisms of urgent and long-term adaptation (Meyerson, Malyshev, 1981). At the same time, the process of urgent adaptation, considered as a temporary measure that the body resorts to in critical situations, has been studied in sufficient detail (Davis, 1960, 1963; Isahakyan, 1972; Tkachenko, 1975; Rohlfs, Daniel, Premont et al., 1995; Beattie, Black, Wood et al., 1996; Marmonier, Duchamp, Cohen-Adad et al., 1997). During this period, the increased production of various signaling factors, including hormonal factors, induces a significant local and systemic restructuring of metabolism in various organs and tissues, which ultimately determines true, long-term adaptation (Khochachka and Somero, 1988). Activation of biosynthetic processes at the level of replication and transcription determines the structural changes that develop in this case, which are manifested by hypertrophy and hyperplasia of cells and organs (Meyerson, 1986). Therefore, the study of the biochemical foundations of adaptation to long-term exposure to disturbing factors is not only of scientific but also of great practical interest, especially from the point of view of the prevalence of maladaptive diseases (Lopez-Torres et al., 1993; Pipkin, 1995; Wallace and Bell, 1995; Sun et al. al., 1996).
Undoubtedly, the development of long-term adaptation of the body is a very complex process, which is realized with the participation of the entire complex of a hierarchically organized system of metabolism regulation, and many aspects of the mechanism of this regulation remain unknown. According to the latest literature data, the adaptation of the body to long-acting disturbing factors begins with local and systemic activation of the phylogenetically most ancient process of free radical oxidation, leading to the formation of physiologically important signaling molecules in the form of reactive oxygen and nitrogen species - nitric oxide, superoxide and hydroxyl. radical, hydrogen peroxide, etc. These metabolites play a leading mediator role in the adaptive local and systemic regulation of metabolism by autocrine and paracrine mechanisms (Sundaresan, Yu, Ferrans et. al., 1995; Finkel, 1998; Givertz, Colucci, 1998).
In this regard, when studying the physiological and pathophysiological aspects of adaptive and maladaptive reactions, the issues of regulation by free-radical metabolites are occupied, and the issues of biochemical mechanisms of adaptation during prolonged exposure to oxidative stress inducers are of particular relevance (Cowan, Langille, 1996; Kemeny, Peakman, 1998; Farrace, Cenni, Tuozzi et al., 1999).
Undoubtedly, the greatest information in this regard can be obtained from experimental studies on the corresponding "models" of common types of oxidative stress. As such, the best known models are exogenous oxidative stress caused by cold exposure and endogenous oxidative stress arising from a deficiency of vitamin E, one of the most important membrane antioxidants. These models were used in this work to elucidate the biochemical foundations of the organism's adaptation to long-term oxidative stress.
In accordance with numerous literature data (Spirichev, Matusis, Bronstein, 1979; Aloia, Raison, 1989; Glofcheski, Borrelli, Stafford, Kruuv, 1993; Beattie, Black, Wood, Trayhurn, 1996), we found that the daily 8-hour cold exposure for 24 weeks led to a pronounced increase in the concentration of malondialdehyde in erythrocytes. This indicates the development of chronic oxidative stress under the influence of cold. Similar changes took place in the body of rats kept for the same period on a diet devoid of vitamin E. This fact is also consistent with the observations of other researchers (Masugi,
Nakamura, 1976; Tamai., Miki, Mino, 1986; Archipenko, Konovalova, Dzhaparidze et al., 1988; Matsuo, Gomi, Dooley, 1992; Cai, Chen, Zhu et al., 1994). However, the causes of oxidative stress in long-term intermittent exposure to cold and oxidative stress in long-term tocopherol deficiency are different. If in the first case the cause of the stress state is the impact of an external factor - cold, which causes an increase in the production of oxyradicals due to the induction of the synthesis of an uncoupling protein in mitochondria (Nohl, 1994; Bhaumik, Srivastava, Selvamurthy et al., 1995; Rohlfs, Daniel, Premont et al. ., 1995; Beattie, Black, Wood et al., 1996; Femandez-Checa, Kaplowitz, Garcia-Ruiz et al., 1997; Marmonier, Duchamp, Cohen-Adad et al., 1997; Rauen, de Groot, 1998 ), then with a deficiency of the membrane antioxidant tocopherol, the cause of oxidative stress was a decrease in the rate of neutralization of oxyradical mediators (Lawler, Cline, He, Coast, 1997; Richter, 1997; Polyak, Xia, Zweier et. al., 1997; Sen, Atalay, Agren et al., 1997; Higashi, Sasaki, Sasaki et al., 1999). Given the fact that prolonged exposure to cold and vitamin E deficiency cause the accumulation of reactive oxygen species, one could expect the transformation of the physiological regulatory role of the latter into a pathological one, with cell damage due to peroxidation of biopolymers. In connection with the generally accepted idea until recently about the damaging effect of reactive oxygen species, cold and tocopherol deficiency are considered as factors provoking the development of many chronic diseases (Cadenas, Rojas, Perez-Campo et al., 1995; de Gritz, 1995; Jain, Wise , 1995; Luoma, Nayha, Sikkila, Hassi., 1995; Barja, Cadenas, Rojas et al., 1996; Dutta-Roy, 1996; Jacob, Burri, 1996; Snircova, Kucharska, Herichova et al., 1996; Va- Squezvivar, Santos, Junqueira, 1996; Cooke, Dzau, 1997; Lauren, Chaudhuri, 1997; Davidge, Ojimba, Mc Laughlin, 1998; Kemeny, Peakman, 1998; Peng, Kimura, Fregly, Phillips, 1998; Nath, Grande, Croatt et al., 1998; Newaz and Nawal, 1998; Taylor, 1998). Obviously, in the light of the concept of the mediator role of reactive oxygen species, the realization of the possibility of transforming physiological oxidative stress into pathological one largely depends on the adaptive increase in the activity of antioxidant enzymes. In accordance with the concept of an antioxidant enzyme complex as a functionally dynamic system, there is a recently discovered phenomenon of substrate induction of gene expression of all three major antioxidant enzymes - superoxide dismutase, catalase, and glutathione peroxidase (Peskin, 1997; Tate, Miceli, Newsome, 1995; Pinkus, Weiner, Daniel, 1996; Watson, Palmer. , Jauniaux et al., 1997; Sugino, Hirosawa-Takamori, Zhong 1998). It is important to note that the effect of such induction has a fairly long lag period, measured in tens of hours and even days (Beattie, Black, Wood, Trayhurn, 1996; Battersby, Moyes, 1998; Lin, Coughlin, Pilch, 1998). Therefore, this phenomenon can lead to an acceleration of the inactivation of reactive oxygen species only under prolonged exposure to stress factors.
The studies carried out in the work showed that long-term intermittent exposure to cold caused a harmonious activation of all the studied antioxidant enzymes. This is consistent with the opinion of Bhaumik G. et al (1995) on the protective role of these enzymes in limiting complications during prolonged cold stress.
At the same time, only superoxide dismutase activation was recorded in the erythrocytes of vitamin E-deficient rats at the end of the 24-week observation period. It should be noted that such an effect was not observed in previous similar studies (Xu, Diplock, 1983; Chow, 1992; Matsuo, Gomi, Dooley, 1992; Walsh, Kennedy, Goodall, Kennedy, 1993; Cai, Chen, Zhu et al. , 1994; Tiidus, Houston, 1994; Ashour, Salem, El Gadban et al., 1999). However, it should be noted that the increase in the activity of superoxide dismutase was not accompanied by an adequate increase in the activity of catalase and glutathione peroxidase and did not prevent the development of the damaging effect of reactive oxygen species. The latter was evidenced by a significant accumulation in erythrocytes of the product of lipid peroxidation - malonidialdehyde. It should be noted that the peroxidation of biopolymers is currently considered as the main cause of pathological changes in avitaminosis E (Chow, Ibrahim, Wei and Chan, 1999).
The effectiveness of antioxidant protection in experiments on the study of cold exposure was evidenced by the absence of pronounced changes in hematological parameters and the preservation of erythrocyte resistance to the action of various hemolytics. Similar results were previously reported by other researchers (Marachev, 1979; Rapoport, 1979; Sun, Cade, Katovich, Fregly, 1999). On the contrary, in animals with E-avitaminosis, a complex of changes was observed that indicated the damaging effect of reactive oxygen species: anemia with intravascular hemolysis, the appearance of erythrocytes with reduced resistance to hemolytics. The latter is considered a very characteristic manifestation of oxidative stress in E-vitaminosis (Brin, Horn, Barker, 1974; Gross, Landaw, Oski, 1977; Machlin, Filipski, Nelson et al., 1977; Siddons, Mills, 1981; Wang, Huang, Chow, 1996). The foregoing convinces of the body's significant capabilities to neutralize the consequences of oxidative stress of external origin, in particular, caused by cold, and the inferiority of adaptation to endogenous oxidative stress in the case of E-avitaminosis.
The group of antioxidant factors in erythrocytes also includes the NADPH generation system, which is a cofactor for heme oxygenase, glutathione reductase, and thioredoxin reductase, which reduce iron, glutathione, and other thio compounds. In our experiments, a very significant increase in the activity of glucose-6-phosphate dehydrogenase in rat erythrocytes was observed both under the influence of cold and with tocopherol deficiency, which was previously observed by other researchers (Kaznacheev, 1977; Ulasevich, Grozina, 1978;
Gonpern, 1979; Kulikov, Lyakhovich, 1980; Landyshev, 1980; Fudge, Stevens, Ballantyne, 1997). This indicates activation in experimental animals of the pentose phosphate shunt, in which NADPH is synthesized.
The mechanism of development of the observed effect becomes clearer in many respects when analyzing changes in carbohydrate metabolism parameters. An increase in glucose uptake by erythrocytes of animals was observed both against the background of oxidative stress caused by cold and during oxidative stress induced by tocopherol deficiency. This was accompanied by a significant activation of membrane hexokinase, the first enzyme of intracellular carbohydrate utilization, which is in good agreement with the data of other researchers (Lyakh, 1974, 1975; Panin, 1978; Ulasevich, Grozina, 1978; Nakamura, Moriya, Murakoshi. et al., 1997; Rodnick , Sidell, 1997). However, the further transformations of glucose-6-phosphate, which is intensively formed in these cases, differed significantly. Upon adaptation to cold, the metabolism of this intermediate increased both in glycolysis (as evidenced by an increase in the activity of hexophosphate isomerase and aldolase) and in the pentose phosphate pathway. The latter was confirmed by an increase in the activity of glucose-6-phosphate dehydrogenase. At the same time, in E-avitaminous animals, the restructuring of carbohydrate metabolism was associated with an increase in the activity of only glucose-6-phosphate dehydrogenase, while the activity of key glycolysis enzymes did not change or even decreased. Therefore, in any case, oxidative stress causes an increase in the rate of glucose metabolism in the pentose phosphate shunt, which ensures the synthesis of NADPH. This seems to be very appropriate in the context of increasing cell demand for redox equivalents, in particular NADPH. It can be assumed that in E-avitaminous animals this phenomenon develops to the detriment of glycolytic energy-producing processes.
The noted difference in the effects of exogenous and endogenous oxidative stress on glycolytic energy production also affected the energy status of cells, as well as energy consumption systems. Under cold exposure, there was a significant increase in the concentration of ATP + ADP with a decrease in the concentration of inorganic phosphate, an increase in the activity of total ATP-ase, Mg-ATP-ase and Na+,K+-ATP-ase. Conversely, in the erythrocytes of rats with E-avitaminosis, a decrease in the content of macroergs and ATPase activity was observed. At the same time, the calculated ATP + ADP / Pn index confirmed the available information that cold, but not E-avitaminous oxidative stress, is characterized by the prevalence of energy production over energy consumption (Marachev, Sorokovoy, Korchev et al., 1983; Rodnick, Sidell, 1997; Hardewig, Van Dijk, Portner, 1998).
Thus, with prolonged intermittent exposure to cold, the restructuring of the processes of energy production and energy consumption in the animal body had a clear anabolic character. This is confirmed by the observed acceleration of the increase in body weight of animals. The disappearance of the hypothermic reaction to cold in rats by the 8th week of the experiment indicates a stable adaptation of their organism to cold and, consequently, the adequacy of adaptive metabolic transformations. At the same time, judging by the main morphofunctional, hematological, and biochemical parameters, changes in energy metabolism in E-avitaminous rats did not lead to an adaptively appropriate result. It seems that the main reason for such an organism's response to tocopherol deficiency is the outflow of glucose from energy-producing processes into the processes of formation of the endogenous antioxidant NADPH. It is likely that the severity of adaptive oxidative stress is a kind of regulator of glucose metabolism in the body: this factor is able to turn on and enhance the production of antioxidants during glucose metabolism, which is more significant for the survival of the body under conditions of a powerful damaging effect of reactive oxygen species than the production of macroergs.
It should be noted that, according to modern data, oxygen radicals are inducers of the synthesis of individual replication and transcription factors that stimulate adaptive proliferation and differentiation of cells in various organs and tissues (Agani and Semenza, 1998). At the same time, one of the most important targets for free radical mediators are transcription factors of the NFkB type, which induce the expression of genes for antioxidant enzymes and other adaptive proteins (Sundaresan, Yu, Ferrans et. al, 1995; Finkel, 1998; Givertz, Colucci, 1998). Thus, one can think that it is this mechanism that is activated during cold-induced oxidative stress and provides an increase in the activity of not only specific antioxidant defense enzymes (superoxide dismutase, catalase, and glutathione peroxidase), but also an increase in the activity of enzymes of the pentose phosphate pathway. With a more pronounced oxidative stress caused by a deficiency of the membrane antioxidant, tocopherol, the adaptive substrate inducibility of these components of the antioxidant defense is realized only partially and, most likely, is not effective enough. It should be noted that the low efficiency of this system ultimately led to the transformation of physiological oxidative stress into pathological.
The data obtained in the work allow us to conclude that the result of adaptive transformations of metabolism in response to disturbing environmental factors, in the development of which reactive oxygen species are involved, is largely determined by the adequacy of the associated increase in the activity of the main antioxidant enzymes, as well as enzymes of the NADPH-generating pentose phosphate pathway. breakdown of glucose. In this regard, when the conditions for the existence of a macroorganism change, especially during the so-called environmental disasters, the severity of oxidative stress and the activity of enzymatic antioxidants should become not only an object of observation, but also one of the criteria for the effectiveness of the body's adaptation.
List of references for dissertation research candidate of biological sciences Skuryatina, Yulia Vladimirovna, 2001
1. Abrarov A.A. Effect of fat and fat-soluble vitamins A, D, E on the biological properties of erythrocytes: Diss. doc. honey. Sciences. M., 1971.- S. 379.
2. Ado A. D., Ado N. A., Bochkov G. V. Pathological physiology.- Tomsk: Publishing House of TSU, 1994.- P. 19.
3. Asatiani V. S. Enzymatic methods of analysis. M.: Nauka, 1969. - 740 p.
4. Benisovich V. I., Idelson L. I. Formation of peroxides and composition of fatty acids in erythrocyte lipids of patients with Marchiafava Micheli’s disease // Probl. hematol. and transfusion, blood. - 1973. - No. 11. - S. 3-11.
5. Bobyrev VN, Voskresensky ON Changes in the activity of antioxidant enzymes in lipid peroxidation syndrome in rabbits // Vopr. honey. chemistry. 1982. - Vol. 28(2). - S. 75-78.
6. Viru A. A. Hormonal mechanisms of adaptation and training. M.: Nauka, 1981.-S. 155.
7. Goldstein D. L., Brown M. S. Genetic aspects of diseases // Internal diseases / Under. ed. E. Braunwald, K. D. Isselbacher, R. G. Petersdorf and others - M .: Medicine, 1993.- T. 2.- P. 135.
8. Datsenko 3. M., Donchenko G. V., Shakhman O. V., Gubchenko K. M., Khmel T. O. The role of phospholipids in the functioning of various cell membranes under conditions of antioxidant system disturbance // Ukr. biochem. j.- 1996.- v. 68(1).- S. 49-54.
9. Yu. Degtyarev V. M., Grigoriev G. P. Automatic recording of acid erythrograms on the EFA-1 densitometer //Lab. case.- 1965.- No. 9.- S. 530-533.
10. P. Derviz G. V., Byalko N. K. Refinement of the method for determining hemoglobin dissolved in blood plasma // Lab. case.- 1966.- No. 8.- S. 461-464.
11. Deryapa N. R., Ryabinin I. F. Human adaptation in the polar regions of the Earth.- L .: Medicine, 1977.- P. 296.
12. Jumaniyazova K. R. Effect of vitamins A, D, E on peripheral blood erythrocytes: Diss. cand. honey. Sciences. - Tashkent, 1970. - S. 134.
13. Donchenko G. V., Metal’nikova N. P., Palivoda O. M. et al. Regulation of ubiquinone and protein biosynthesis in rat liver with E-hypovitaminosis by a-tocopherol and actinomycin D, Ukr. biochem. J.- 1981.- T. 53(5).- S. 69-72.
14. Dubinina E. E., Salnikova L. A., Efimova L. F. Activity and isoenzyme spectrum of erythrocyte and plasma superoxide dismutase // Lab. case.- 1983.-№10.-S. 30-33.
15. Isahakyan JI. A. Metabolic structure of temperature adaptations D.: Nauka, 1972.-S. 136.
16. Kaznacheev V.P. Biosystem and adaptation // Report at the II session of the Scientific Council of the USSR Academy of Sciences on the problem of applied human physiology. - Novosibirsk, 1973.-S. 74.
17. Kaznacheev V.P. Problems of human adaptation (results and prospects) // 2nd All-Union. conf. on adaptation of the person to various. geographical, climatic, and industrial conditions: abstract. dokl.- Novosibirsk, 1977.- v. 1.-S. 3-11.
18. Kaznacheev V.P. Modern aspects of adaptation. - Novosibirsk: Nauka, 1980.-S. 191.
19. Kalashnikov Yu. K., Geisler B. V. On the method of determining blood hemoglobin using acetone cyanohydrin // Lab. case.- 1975.- No. 6.- SG373-374.
20. Kandror I. S. Essays on human physiology and hygiene in the Far North. - M .: Medicine, 1968. - P. 288.
21. Kashevnik L. D. Metabolism in beriberi S.- Tomsk., 1955.- S. 76.
22. Korovkin B. F. Enzymes in the diagnosis of myocardial infarction.- L: Nauka, 1965.- P. 33.
23. Kulikov V. Yu., Lyakhovich V. V. Reactions of free radical oxidation of lipids and some indicators of oxygen metabolism // Mechanisms of human adaptation in high latitudes / Ed. V. P. Kaznacheeva.- L .: Medicine, 1980.- S. 60-86.
24. Landyshev S.S. Adaptation of erythrocyte metabolism to the action of low temperatures and respiratory failure // Adaptation of humans and animals in various climatic zones / Ed. M. 3. Zhits.- Chita, 1980.- S. 51-53.
25. Lankin V. Z., Gurevich S. M., Koshelevtseva N. P. The role of lipid peroxides in the pathogenesis of atherosclerosis. Detoxification of lipoperoxides by the glutathione peroxidase system in the aorta // Vopr. honey. Chemistry. - 1976. - No. 3, - S. 392-395.
26. Lyakh L.A. On the stages of formation of adaptation to cold // Theoretical and practical problems of the effect of low temperatures on the body: Tez. IV All-Union. Conf.- 1975.- S. 117-118.
27. Marachev A. G., Sorokovoi V. I., Korchev A. V. et al. Bioenergetics of erythrocytes in the inhabitants of the North // Human Physiology.- 1983.- No. 3.- P. 407-415.
28. Marachev A.G. Structure and function of human erythron in the conditions of the North // Biological problems of the North. VII symposium. Human adaptation to the conditions of the North / Ed. V.F. Burkhanova, N.R. Deryapy.- Kirovsk, 1979.- S. 7173.
29. Matusis I. I. Functional relationships of vitamins E and K in the metabolism of the animal organism // Vitamins.- Kyiv: Naukova Dumka, 1975.- Vol. 8.-S. 71-79.
30. Meyerson F. 3., Malyshev Yu. I. The phenomenon of adaptation and stabilization of structures and protection of the heart.- M: Medicine, 1981.- P. 158.
31. Meyerson F. 3. Basic patterns of individual adaptation // Physiology of adaptation processes. M.: Nauka, 1986.- S. 10-76.
32. Panin JI. E. Some biochemical problems of adaptation // Medico-biological aspects of adaptation processes / Ed. J.I. P. Nepomnyashchikh.-Novosibirsk.: Science.-1975a.-S. 34-45.
33. Panin L. E. The role of hormones of the pituitary-adrenal system and pancreas in the violation of cholesterol metabolism in some extreme conditions: Diss. doc. honey. nauk.- M., 19756.- S. 368.
34. Panin L. E. Energy aspects of adaptation. - L.: Medicine, 1978. - 192 p. 43. Panin L. E. Features of energy metabolism // Mechanisms of human adaptation to high latitude conditions / Ed. V. P. Kaznacheeva.- L .: Medicine, 1980.- S. 98-108.
35. Peskin A. V. Interaction of active oxygen with DNA (Review) // Biochemistry.- 1997.- T. 62.- No. 12.- P. 1571-1578.
36. Poberezkina N. B., Khmelevsky Yu. V. Disturbances in the structure and function of erythrocyte membranes E in beriberi rats and its correction with antioxidants // Ukr. biochem. j.- 1990.- v. 62(6).- S. 105-108.
37. Pokrovsky A. A., Orlova T. A., Pozdnyakov A. JL Influence of tocopherol deficiency on the activity of certain enzymes and their isoenzymes in the testes of rats // Vitamins and reactivity of the organism: Proceedings of the MOIP.- M., 1978.-T. 54.- S. 102-111.
38. Rapoport Zh. Zh. Adaptation of the child in the North.- L .: Medicine, 1979.- P. 191.
39. Rossomahin Yu. I. Features of thermoregulation and body resistance to contrasting effects of heat and cold under different modes of temperature adaptation: Abstract of the thesis. diss. cand. biol. Sciences.- Donetsk, 1974.- S. 28.
40. Seits, I.F., On the quantitative determination of adenosine tri- and adenosine diphosphates, Byull. exp. biol. and medical - 1957. - No. 2. - S. 119-122.
41. Sen IP Development of E-vitamin deficiency in white rats fed qualitatively different fats: Diss. cand. honey. nauk.- M., 1966.- S. 244.
42. Slonim, A.D., Physiological mechanisms of natural adaptations of animals and humans, Dokl. for the yearly session Academic Council dedicated. memory of acad. K. M. Bykova. - JL, 1964.
43. Slonim A. D. Physiological adaptations and peripheral structure of reflex responses of the organism // Physiological adaptations to heat and cold / Ed. A. D. Slonim.- JL: Science, 1969.- S. 5-19.
44. Spirichev V. B., Matusis I. I., Bronstein JL M. Vitamin E. // In the book: Experimental vitaminology / Ed. Yu. M. Ostrovsky.- Minsk: Science and Technology, 1979.- S. 18-57.
45. Stabrovsky E. M. Energy metabolism of carbohydrates and its endocrine regulation under the influence of low environmental temperature on the body: Avto-ref. diss. doc. biol. nauk.- JL, 1975.- S. 44.
46. Tepliy D. JL, Ibragimov F. Kh. Changes in the permeability of erythrocyte membranes in rodents under the influence of fish oil, vitamin E and fatty acids // J. Evolution. Biochemistry and Physiology.- 1975.- v. 11(1).- S. 58-64.
47. Terskov I. A., Gitelzon I. I. Erythrograms as a method of clinical blood testing.- Krasnoyarsk, 1959.- P. 247.
48. Terskov I. A., Gitelzon I. I. The value of dispersion methods for the analysis of erythrocytes in normal and pathological conditions // Questions of biophysics, biochemistry and pathology of erythrocytes.- M .: Nauka, 1967.- P. 41-48.
49. Tkachenko E. Ya. On the ratio of contractile and non-contractile thermogenesis in the body during adaptation to cold // Physiological adaptation to cold, mountain and subarctic conditions / Ed. K. P. Ivanova, A. D. Slonim.-Novosibirsk: Nauka, 1975.- P. 6-9.
50. Uzbekov G. A., Uzbekov M. G. Highly sensitive micromethod for photometric determination of phosphorus // Lab. case.- 1964.- No. 6.- S. 349-352.
51. Khochachka P., Somero J. Biochemical adaptation: Per. from English. M.: Mir, 1988.-576 p.
52. Shcheglova, AI, Adaptive Changes in Gas Exchange in Rodents with Different Ecological Specializations, Physiological Adaptations to Heat and Cold, Ed. A. D. Slonim.- L.: Nauka, 1969.- S. 57-69.
53. Yakusheva I. Ya., Orlova LI Method for determination of adenosine triphosphatases in hemolysates of blood erythrocytes // Lab. case.- 1970.- No. 8.- S. 497-501.
54. Agani F., Semenza G. L. Mersalyl is a novel inducer of vascular endothelial growth factor gene expression and hypoxia-inducible factor 1 activity // Mol. Pharmacol.- 1998.- Vol. 54(5).-P. 749-754.
55. Ahuja B. S., Nath R. A kinetik study of superoxide dismutase in normal human erytrocytes and its possible role in anemia and radiation damage // Simpos. on control in mechanisms cell, processes. - Bombey, 1973. - P. 531-544.
56. Aloia R. C., Raison J. K. Membrane function in mammalian hibernation // Bio-chim. Biophys. Acta.- 1989.- Vol. 988.- P. 123-146.
57. Asfour R. Y., Firzli S. Hematologic stadies in undernourished children with low serum vitamin E levels // Amer. J.Clin. Nutr.- 1965.- Vol. 17(3).-P. 158-163.
58. Ashour M. N., Salem S. I., El Gadban H. M., Elwan N. M., Basu T. K. Antioxidant status in children with protein-energy malnutrition (PEM) living in Cairo, Egypt // Eur. J.Clin. Nutr.- 1999.- Vol. 53(8).-P. 669-673.
59. Bang H. O., Dierberg J., Nielsen A. B. Plasma lipid and lipoprotein pattern in Greenlandic west coast Eskimos // Lancet.- 1971.- Vol. 7710(1).-P. 1143-1145.
60. Barja G., Cadenas S., Rojas C., et al. Effect of dietary vitamin E levels on fatty acid profiles and nonenzymatic lipid peroxidation in the guinea pig liver // Lipids.-1996.- Vol. 31(9).-P. 963-970.
61. Barker M. O., Brin M. Mechanisms of lipid peroxidation in erythrocytes of vitamin E deficients rats and in phospholipid model sistems // Arch. Biochem. and Biophys.- 1975.- Vol. 166(1).-P. 32-40.
62. Battersby B. J., Moyes C. D. Influence of acclimation temperature on mitochondrial dna, rna and enzymes in skeletal muscle // APStracts.- 1998.- Vol. 5.- P. 195.
63. Beattie J. H., Black D. J., Wood A. M., Trayhurn P. Cold-induced expression of the metallothionein-1 gene in brown adipose tissue of rats, Am. J. Physiol.-1996.-Vol. 270(5).- Pt 2.- P. 971-977.
64. Bhaumik G., Srivastava K. K., Selvamurthy W., Purkayastha S. S. The role of free radicals in cold injuries // Int. J. Biometeorol.- 1995.- Vol. 38(4).-P. 171-175.
65. Brin M., Horn L. R., Barker M. O. Relationship between fatty acid composition oferithrocytes and susceptibility to vitamin E deficiency // Amer. J.Clin. Nutr.-%1974.-Vol. 27(9).-P. 945-950.
66. Caasi P. I., Hauswirt J. W., Nair P. P. Biosynthesis of heme in vitamin E deficiency // Ann. N. Y. Acad. Sci.- 1972.- Vol. 203.- P. 93-100.
67. Cadenas S., Rojas C., Perez-Campo R., Lopez-Torres M., Barja G. Vitamin E protects guinea pig liver from lipid peroxidation without depressing levels of antioxidants//Int. J Biochem. cell. Biol.- 1995.-Vol. 27(11).-P. 1175-1181.
68 Cai Q. Y., Chen X. S., Zhu L. Z., et al. Biochemical and morphological changes in the lenses of selenium and/or vitamin E deficient rats // Biomed. Environ. Sci.-1994.-Vol. 7(2).-P. 109-115.
69. Cannon R. O. Role of nitric oxide in cardiovascular disease: focus on the endothelium // Clin. Chem.- 1998.- Vol. 44.- P. 1809-1819.
70. Chaudiere J., Clement M., Gerard D., Bourre J. M. Brain alterations induced by vitamin E deficiency and intoxication with methyl ethyl ketone peroxide // Neuro-toxicology.- 1988.- Vol. 9(2).-P. 173-179.
71. Chow C. K. Distribution of tocopherols in human plasma and red blood cells // Amer. J.Clin. Nutr.- 1975.- Vol. 28(7).-P. 756-760.
72. Chow C. K. Oxidative damage in the red cells of vitamin E-deficient rats // Free. Radic. Res. Commun.- 1992 vol. 16(4).-P. 247-258.
73. Chow C. K., Ibrahim W., Wei Z., Chan A. C. Vitamin E regulates mitochondrial hydrogen peroxide generation // Free Radic. Biol. Med.- 1999.- Vol. 27 (5-6).- P. 580-587.
74. Combs G. F. Influences of dietary vitamin E and selenium on the oxidant defense system of the chick//Poult. Sci.- 1981.- Vol. 60(9).- P. 2098-2105.
75. Cooke J. P., Dzau V. J. Nitric oxide synthase: Role in the Genesis of Vascular Disease // Ann. Rev. Med.- 1997.- Vol. 48.- P. 489-509.
76. Cowan D. B., Langille B. L. Cellular and molecular biology of vascular remodeling // Current Opinion in Lipidology.- 1996.- Vol. 7.- P. 94-100.
77. Das K. S., Lewis-Molock Y., White C. W. Elevation of manganese superoxide dismutase gene expression by thioredoxin, Am. J. Respir. Cell Mol. Biol.- 1997.-Vol. 17(6).-P. 12713-12726.
78. Davidge S. T., Ojimba J., McLaughlin M. K. Vascular Function in the Vitamin E Deprived Rat. An Interaction Between Nitric Oxide and Superoxide Anions // Hypertension.- 1998.- Vol. 31.- P. 830-835.
79. Davis T. R. A. Shivering and nonshivering heat production in animals and man, Cold Injury: Ed. S. H. Horvath.- N. Y., I960.- P. 223-269.
80. Davis T. R. A. Nonshivering thermogenesis, Feder. Proc.- 1963.- Vol. 22(3).-P. 777-782.
81. Depocas F. Calorigenesis from various organ systems in the whole animal // Feder. Proc.-I960.-Vol. 19(2).-P. 19-24.
82. Desaultes M., Zaror-Behrens G., Hims-Hagen J. Increased purine nucleotide binding, altered polipeptide composition and thermogenesis in brown adipose tissue mitochondria of cold-acclimated rats // Can. J. Biochem.- 1978.- Vol. 78(6).-P. 378-383.
83. Drexler H., Hornig B. Endothelial dysfunction in human disease // J. Mol. cell. Cardiol.- 1999.- Vol. 31(1).-P. 51-60.
84. Dutta-Roy A. K. Therapy and clinical trials // Current Opinion in Lipidology.-1996.-Vol. 7.-P. 34-37.
85. Elmadfa I., Both-Bedenbender N., Sierakowski B., Steinhagen-Thiessen E. Significance of vitamin E in aging // Z. Gerontol.- 1986.- Vol. 19(3).-P. 206-214.
86. Farrace S., Cenni P., Tuozzi G., et al. Endocrine and psychophysiological aspects of human adaptation to the extreme //Physiol. Behav.- 1999.- Vol.66(4).- P.613-620.
87. Fernandez-Checa, J. C., Kaplowitz N., Garcia-Ruiz C., et al. Importance and characteristics of glutahione transport in mitochondria: defense against TNF-induced oxidative stress and defect induced by alcohol // APStracts.- 1997.-Vol.4.- P. 0073G.
88. Finkel T. Oxygen radicals and signaling // Current Opinion in Cell Biology.-1998.- Vol. 10.-p. 248-253.
89. Photobiol.- 1993.- Vol. 58(2).-P. 304-312.
90. Fudge D. S., Stevens E. D., Ballantyne J. S. Enzyme adaptation along a hetero-thermic tissue the visceral retia mirabilia of the bluefin tuna // APStracts.- 1997.-Vol. 4, - P. 0059R.
91. Givertz M. M., Colucci W. S. New targets for heart-failure therapy: endothelin, inflammatory cytokines, and oxidative stress // Lancet.- 1998.- Vol.352- Suppl 1.-P. 34-38.
92. Glofcheski D. J., Borrelli M. J., Stafford D. M., Kruuv J. Induction of tolerance to hypothermia and hyperthermia by a common mechanism in mammalian cells // J. Cell. Physiol.- 1993.- Vol. 156.- P. 104-111.
93. Chemical Biology.- 1999.- Vol. 3.- P. 226-235.1 ll. Guarnieri C., Flamigni F., Caldarera R. C:, Ferrari R. Myocardial mitochondrial functions in alpha-tocopherol-deficient and -refed rabbits // Adv. Myocardiol.-1982.-Vol.3.-P. 621-627.
94. Hardewig I., Van Dijk P. L. M., Portner H. O. High energy turnover at low temperatures: recovery from exhaustive exercise in antarctic and temperate eelpouts (zoarcidae) // APStracts.- 1998.- Vol. 5.- P. 0083R.
95. Hassan H., Hashins A., van Italie T. B., Sebrell W. H. Syndrom in premature infants anemia associated with low plasma vitamin E level and high poliunsaturated fatty acid diet // Amer. J.Clin. Nutr.-1966.-Vol. 19(3).-P. 147-153.
96. Hauswirth G. W., Nair P. P. Some aspects of vitamin E in expression of biological information, Ann. N. Y. Acad. Sci.- 1972.- Vol. 203.- P. 111-122.
97. Henle E. S., Linn S. Formation, prevention, and repair of DNA damage by iron/hydrogen peroxide // J. Biol, chem.- 1997.- Vol. 272(31).- P. 19095-19098.
98. Higashi Y., Sasaki S., Sasaki N., et al. Daily aerobic exercise improves reactive hyperemia in patients with essential hypertension // Hypertension.- 1999.- Vol. 33(1).-Pt 2.-P. 591-597.
99. Howarth P. H Pathogenic mechanisms: a rational basis for treatment // V. M. J.-1998.-Vol. 316.-p. 758-761.
100. Hubbell R. B., Mendel L. B., Wakeman A. J. A new salt mixture for use in experimental diets // J. Nutr.- 1937.- Vol. 14.- P. 273-285.
101. Jacob R. A., Burri B. J. Oxidative damage and defense // Am. J.Clin. Nutr.-1996.-Vol. 63.- P. 985S-990S.
102. Jain S. K., Wise R. Relationship between elevated lipid peroxides, vitamin E deficiency and hypertension in preeclampsia, Mol. cell. Biochem.- 1995.- Vol. 151(1).-P. 33-38.
103. Karel P., Palkovits M., Yadid G., et al. Heterogeneous neurochemical responses to different stressors: a test of selye's doctrine of nonspecificity // APStracts.-1998.-Vol. 5.-P. 0221R.
104. Kausalya S., Nath J. Interactive role of nitric oxide and superoxide anion in neu-trophil-mediated endothelial cell in injury // J. Leukoc. Biol.- 1998.- Vol. 64(2).-P. 185-191.
105. Kemeny M., Peakman M. Immunology // B. M. J. - 1998.- Vol. 316.- P. 600-603.
106. Kozyreva T. V., Tkachenko E. Y., Kozaruk V. P., Latysheva T. V., Gilinsky M. A. The effects of slow and rapid cooling on catecholamine concentration in arterial plasma and the skin // APStracts.- 1999.- Vol. 6.- P. 0081R.
107. Lauren N., Chaudhuri G. Estrogens and atherosclerosis, Ann. Rev. Pharmacol. Toxicol.- 1997.- Vol. 37.- P. 477-515.
108. Lawler J. M., Cline C. C., Hu Z., Coast J. R. Effect of oxidative stress and acidosis on diaphragm contractile function // Am. J. Physiol.- 1997.- Vol. 273(2).-Pt 2.-P. 630-636.
109. Lin B., Coughlin S., Pilch P. F. Bi-directional regulation of uncoupling protein-3 and glut4 mrna in skeletal muscle by cold // APStracts.- 1998.- Vol. 5.- P. 0115E.
110. Lindquist J. M., Rehnmark S. Ambient temperature regulation of apoptosis in brown adipose tissue // J. Biol. Chem.- 1998.- Vol. 273(46).-P. 30147-30156.
111. Lowry O. H., Rosenbrough N. G., Farr A. L., Randell R. I. Protein measurement with the Folin phenol reagent // J. Biol. Chem.-195L-Vol. 193.- P. 265-275.
112. Luoma P. V., Nayha S., Sikkila K., Hassi J. High serum alpha-tocopherol, albumin, selenium and cholesterol, and low mortality from coronary heart disease in northern Finland//J.Intern. Med.- 1995.-Vol. 237(1).-P. 49-54.
113. Luscher T. F., Noll G., Vanhoutte P. M. Endothelial dysfunction in hypertension // J. Hypertens.- 1996.- Vol. 14(5).-P. 383-393.
114. Machlin L. J., Filipski R., Nelson J., Horn L. R., Brin M. Effect of progressive vitamin E deficiency in the rat // J. Nutr.- 1977.- Vol. 107(7).-P. 1200-1208.
115. Marmonier F., Duchamp C., Cohen-Adad F., Eldershaw T. P. D., Barra H. Hormonal control of thermogenesis in perfused muscle of muscovy ducklings // AP-Stracts.-1997.- Vol. 4.- P. 0286R.
116. Marvin H. N. Erythrocyte survival of rat deficient in vitamin E or vitamin B6 // J. Nutr.- 1963.-Vol. 80(2).-P. 185-190.
117. Masugi F., Nakamura T. Effect of vitamin E deficiency on the level of superoxide dismutase, glutathione peroxidase, catalase and lipid peroxide in rat liver, Int. J. Vitam. Nutr. Res.- 1976.- Vol. 46(2).-P. 187-191.
118. Matsuo M., Gomi F., Dooley M. M. Age-related alterations in antioxidant capacity and lipid peroxidation in brain, liver, and lung homogenates of normal and vitamin E-deficient rats // Mech. Ageing Dev.- 1992.- Vol. 64(3).-P. 273-292.
119. Mazor D., Brill G., Shorer Z., Moses S., Meyerstein N. Oxidative damage in red blood cells of vitamin E deficient patients // Clin. Chim. Acta.- 1997.- Vol. 265(l).-P. 131-137.
120. Mircevova L. The role of Mg++-ATPase (actomyosine-like protein) in maintaining the biconcave shape of erythrocytes // Blut.- 1977.- vol 35(4).- P. 323-327.
121. Mircevova L., Victora L., Kodicek M., Rehackova H., Simonova A. The role of spectrin dependent ATPase in erytrocyte shape maintenance // Biomed. biochim. Acta.- 1983.- Vol. 42(11/12).- P. 67-71.
122. Nair P. P. Vitamin E and metabolic regulation // Ann. N. Y. Acad. Sci.-1972a.-Vol. 203.- P. 53-61.
123. Nair P. P. Vitamine E regulation of the biosintesis of porphirins and heme // J. Agr. and Food Chem.- 1972b.- Vol. 20(3).-P. 476-480.
124. Nakamura T., Moriya M., Murakoshi N., Shimizu Y., Nishimura M. Effects of phenylalanine and tyrosine on cold acclimation in mice // Nippon Yakurigaku Zasshi.-1997.-Vol. 110(1).-P. 177-182.
125. Nath K. A., Grande J., Croatt A., et al. Redox regulation of renal DNA synthesis, transforming growth factor-betal and collagen gene expression // Kidney Int.-1998.- Vol. 53(2).-P. 367-381.
126. Nathan C. Perspectives Series: Nitric Oxide and Nitric Oxide Synthases Inducible Nitric Oxide Synthase: What Difference Does It Make? // J. Clin. Invest.1997.- Vol. 100(10).-P. 2417-2423.
127. Newaz M. A., Nawal N. N. Effect of alpha-tocopherol on lipid peroxidation and total antioxidant status in spontaneously hypertensive rats // Am J Hypertens.1998.-Vol. 11(12).-P. 1480-1485.
128. Nishiyama H., Itoh K., Kaneko Y., et al. Glycine-rich RNA-binding Protein Mediating Cold-inducible Suppression of Mammalian Cell Growth // J. Cell. Biol.- 1997.- Vol. 137(4).-P. 899-908.
129. Nohl H. Generation of superoxide radicals as byproduct of cellular respiration, Ann. Biol. Clin. (Paris).- 1994.- Vol. 52(3).-P. 199-204.
130. Pendergast D. R., Krasney J. A., De Roberts D. Effects of immersio in cool water on lung-exhaled nitric oxide at rest and during exercise // Respir. Physiol.-1999.-Vol. 115(1).-P. 73-81.
131. Peng J. F., Kimura B., Fregly M., Phillips M. I. Reduction of cold-induced hypertension by antisense oligodeoxynucleotides to angiotensinogen mRNA and ATi receptor mRNA in brain and blood // Hypertension.- 1998.- Vol. 31.- P. 13171323.
132. Pinkus R., Weiner L. M., Daniel V. Role of oxidants and antioxidants in the induction of AP-1, NF-kappa B and glutathione S~transferase gene expression // J. Biol. Client.- 1996.- Vol. 271(23).- P. 13422-13429.
133. Pipkin F. B. Fortnightly Review: The hypertensive disorders of pregnancy // BMJ.- 1995.-Vol. 311.-P. 609-613.
134. Reis S. E., Blumenthal R. S., Gloth S. T., Gerstenblith R. G., Brinken J. A. Estrogen acutely abolishes cold-induced coronary vasoconstriction in postmenopausal women // Circulation.- 1994.- Vol. 90.- P. 457.
135. Salminen A., Kainulainen H., Arstila A. U., Vihko V. Vitamin E deficiency and the susceptibility to lipid peroxidation of mouse cardiac and skeletal muscles // Acta Physiol. Scand.- 1984.- Vol. 122(4).-P. 565-570.
136. Sampson G. M. A., Muller D. P. Studies on the neurobiology of vitamin E (al-pha-tocopherol) and some other antioxidant systems in the rat // Neuropathol. Appl. Neurobiol.- 1987.- Vol. 13(4).-P. 289-296.
137. Sen C. K., Atalay M., Agren J., Laaksonen DE, Roy S., Hanninen O. Fish oil and vitamin E supplementation in oxidative stress at rest and after physical exercise // APStracts.- 1997.- Vol. . 4.- P. 0101 A.
138. Shapiro S. S., Mott D. D., Machlin L. J. Altered binding of glyceraldehyde 3 -phosphate dehydrogenase to its binding site in vitamine E - deficient red blood cells // Nutr. Rept. Int.- 1982.- Vol. 25(3).-P. 507-517.
139. Sharmanov A. T., Aidarkhanov V. V., Kurmangalinov S. M. Effect of vitamin E deficiency on oxidative metabolism and antioxidant enzyme activity of macrophages // Ann. Nutr. Metab.- 1990.- Vol. 34(3).-P. 143-146.
140. Siddons R. C., Mills C. F. Glutatione peroxidase activity and erythrocyte stability in calves differing in selenium and vitamin E status, Brit. J. Nutr.-1981.-Vol. 46(2).-P. 345-355.
141. Simonoff M., Sergeant C., Gamier N., et al. Antioxidant status (selenium, vitamins A and E) and aging // EXS.- 1992.- Vol. 62.- P. 368-397.
142. Sklan D., Rabinowitch H. D., Donaghue S. Superoxide dismutase: effect of vitamins A and E, Nutr. Rept. Int.- 1981.- Vol. 24(3).-P. 551-555.
143. Smith S. C., Guilbert L. J., Yui J., Baker P. N., Davidge S. T. The role of reactive nitrogen/oxygen intermediates in cytokine-induced trophoblast apoptosis // Placenta.- 1999.- Vol. 20(4).-P. 309-315.
144. Snircova M., Kucharska J., Herichova I., Bada V., Gvozdjakova A. The effect of an alpha-tocopherol analog, MDL 73404, on myocardial bioenergetics // Bratisl Lek Listy.- 1996.- Vol. 97. P. 355-359.
145. Soliman M. K. Uber die Blutveranderungen bei Ratten nach verfuttem einer Tocopherol und Ubichinon Mangeldiat. 1. Zytologische und biochemische Veranderungen im Blut von vitamin E Mangelratten // Zbl. Veterinarmed.- 1973.-Vol. 20(8).-P. 624-630.
146. Stampfer M. J., Hennekens C. H., Manson J. E., et al. Vitamin E consumption and the risk of coronary disease in women // N. Engl. J. Med.- 1993.- Vol. 328.- P. 1444-1449.
147. Sun J. Z., Tang X. L., Park S. W., et al. Evidence for an Essential Role of Reactive Oxygen Species in the Genesis of Late Preconditioning Against Myocardial Stunning in Conscious Pigs // J. Clin. Invest. 1996, Vol. 97(2).-P. 562-576.
148. Sun Z., Cade J. R., Fregly M. J. Cold-induced hypertension. A model of miner-alocorticoid-induced hypertension// Ann.N.Y.Acad.Sci.- 1997.- Vol.813.- P.682-688.
149. Sun Z., Cade R, Katovich M. J., Fregly M. J. Body fluid distribution in rats with cold-induced hypertension // Physiol. Behav.- 1999.- Vol. 65(4-5).-P. 879-884.
150. Sundaresan M., Yu Z.-X., Ferrans VJ, Irani K., Finkel T. Requirement for generation of H202 for platelet-derived growth factor signal transduction // Science (Wash. DC).- 1995.- Vol . 270.- P. 296-299.
151. Suzuki J., Gao M., Ohinata H., Kuroshima A., Koyama T. Chronic cold exposure stimulates microvascular remodeling preferentially in oxidative muscles in rats // Jpn. J. Physiol.- 1997.- Vol. 47(6).-P. 513-520.
152. Tamai H., Miki M., Mino M. Hemolysis and membrane lipid changes induced by xanthine oxidase in vitamin E deficient red cells // J. Free Radic. Biol. Med.-1986.-Vol. 2(1).-P. 49-56.
153. Tanaka M., Sotomatsu A., Hirai S. Aging of the brain and vitamin E // J. Nutr. sci. Vitaminol. (Tokyo).- 1992.- Spec. No.-P. 240-243.
154. Tappel, A. L. Free radical lipid peroxidation damage and its inhibition by vitamin E and selenium, Fed. Proc.- 1965.- Vol. 24(1).-P. 73-78.
155. Tappel, A. L. Lipid peroxidation damage to cell components, Fed. Proc.- 1973.-Vol. 32(8).-P. 1870-1874.
156. Taylor A.J. N. Asthma and allergy // B. M. J.- 1998.- Vol. 316.- P. 997-999.
157. Tate D. J., Miceli M. V., Newsome D. A. Phagocytosis and H2C>2 induce catalase and metaliothionein irene expression in human retinal pigment epithelial cells // Invest. Onithalmol. Vis. Sci.- 1995.- Vol. 36.- P. 1271-1279.
158. Tensuo N. Effect of daily infusion of noradrenaline on metabolism and skin temperature in rabbits // J. Appl. Physiol.- 1972.- Vol. 32(2).-P. 199-202.
159. Tiidus P. M., Houston M. E. Antioxidant and oxidative enzyme adaptations to vitamin E deprivation and training // Med. sci. sports. Exerc.- 1994.- Vol. 26(3).-P. 354-359.
160. Tsen C. C., Collier H. B. The protective action of tocopherol against hemolisis of rat eritrocites by dialuric acid // Canada. J Biochem. Physiol.-I960.-Vol. 38(9).-P. 957-964.
161. Tudhope G. R., Hopkins J. Lipid peroxidation in human erythrocytes in tocopherol deficiency // Acta Haematol.- 1975.- Vol. 53(2).-P. 98-104.
162. Valentine J. S., Wertz D. L., Lyons T. J., Liou L.-L., Goto J. J., Gralla E. B. The dark side of dioxygen biochemistry // Current Opinion in Chemical Biology.-1998.-Vol. 2.-P. 253-262.
163. Vransky V. K. Red blood cell membrane resistanse // Biophys. Membrane Transport.- Wroclaw.- 1976.- Part 2.- P. 185-213.
164. Vuillanine R. Role biologiqe et mode d" action des vitamins E // Rec. med vet.-1974.-Vol. 150(7).-P. 587-592.
165. Wang J., Huang C. J., Chow C. K. Red cell vitamin E and oxidative damage: a dual role of reducing agents, Free Radic. Res.- 1996 Vol. 24(4).-P. 291-298.
166. Wagner B. A., Buettner G. R., Burns C. P. Vitamin E slows the rate of free radical-mediated lipid peroxidation in cells // Arch. Biochem. Biophys.- 1996.- Vol. 334.-p. 261-267.
167. Wallace J. L., Bell C. J. Gastroduodenal mucosal defense // Current Opinion in Gastroenterology 1994 .-Vol. 10.-p. 589-594.
168. Walsh D. M., Kennedy D. G., Goodall E. A., Kennedy S. Antioxidant enzyme activity in the muscles of calves depleted of vitamin E or selenium or both // Br. J. Nutr.- 1993.- Vol. 70(2).-P. 621-630.
169. Watson A. L., Palmer M. E., Jauniaux E., Burton G. J. Variations in expression of copper/zinc superoxide dismutase in villous trophoblast of the human placenta with gestational age // Placenta.- 1997.- Vol. 18(4).-P. 295-299.
170. Young J. B., Shimano Y. Effects of rearing temperature on body weight and abdominal fat in male and female rats // APStracts.-1991.- Vol. 4.- P. 041 OR.
171. Zeiher A. M., Drexler H., Wollschlager H., Just H. Endothelial dysfunction of the coronary microvasculature is associated with coronary blood flow regulation in patients with early atherosclerosis // Circulation.- 1991.- Vol. 84.- P. 19841992.
Please note that the scientific texts presented above are posted for review and obtained through recognition of the original texts of dissertations (OCR). In this connection, they may contain errors related to the imperfection of recognition algorithms. There are no such errors in the PDF files of dissertations and abstracts that we deliver.
The ability to adapt to cold is determined by the magnitude of the body's energy and plastic resources; in their absence, adaptation to cold is impossible. The response to cold develops in stages and in almost all body systems. The early stage of adaptation to cold can be formed at a temperature of 3C about within 2 minutes, and at 10C about for 7 minutes.
From the side of the cardiovascular system, 3 phases of adaptive reactions can be distinguished. The first 2 are optimal (desirable) when exposed to cold for the purpose of hardening. They are manifested in the inclusion, through the nervous and endocrine systems, of the mechanisms of non-contractile thermogenesis, against the background of narrowing of the vascular bed in the skin, resulting in heat production and an increase in the temperature of the "core", which leads to a reflex increase in blood flow in the skin and increased heat transfer, including through inclusion of reserve capillaries. Outwardly, it looks like a uniform hyperemia of the skin, a pleasant feeling of warmth and cheerfulness.
The third phase develops when overloaded with a cold agent in terms of intensity or duration. Active hyperemia is replaced by passive (congestive), blood flow slows down, the skin acquires a bluish tint (venous congestive hyperemia), muscle tremor appears, "goosebumps". This response phase is not desirable. It indicates the depletion of the body's compensatory capabilities, their insufficiency to compensate for heat loss and the transition to contractile thermogenesis.
The reactions of the cardiovascular system are formed not only from the redistribution of blood flow in the skin depot. Cardiac activity slows down, the ejection fraction becomes larger. There is a slight decrease in blood viscosity and an increase in blood pressure. With an overdose of the factor (third phase), an increase in blood viscosity occurs with a compensatory movement of the interstitial fluid into the vessels, which leads to tissue dehydration.
Breathing regulation
Under normal conditions, respiration is regulated by the deviation of the partial pressure of O 2 and CO 2 and the pH value in arterial blood. Moderate hypothermia excites the respiratory centers and depresses the pH-sensitive chemoreceptors. With prolonged cold, a spasm of the bronchial muscles joins, which increases the resistance to breathing and gas exchange, and also reduces the chemosensitivity of the receptors. The ongoing processes underlie cold hypoxia, and in case of failure of adaptation to the so-called "polar" shortness of breath. The respiratory organs react to therapeutic cold procedures with a delay at the first moment, followed by an increase for a short time. In the future, breathing slows down and becomes deep. There is an increase in gas exchange, oxidative processes, and basal metabolism.
metabolic reactions
Metabolic reactions cover all aspects of the exchange. The main direction, of course, is to increase heat production. First of all, non-shivering thermogenesis is activated by mobilizing lipid metabolism (the concentration of free fatty acids in the blood increases by 300% under the influence of cold) and carbohydrates. The tissue consumption of oxygen, vitamins, macro- and microelements is also activated. In the future, with uncompensated heat losses, shivering thermogenesis is switched on. The thermogenic activity of shivering is higher than that during the production of voluntary contractile movements, because. no work is done, and all energy is converted into heat. All muscles are included in this reaction, even the respiratory muscles of the chest.
Water-salt exchange
In the case of acute exposure to cold, the sympathetic-adrenal system is initially activated and the secretion of the thyroid gland increases. Increased production of antidiuretic hormone, which reduces sodium reabsorption in the renal tubules and increases fluid excretion. This leads to the development of dehydration, hemoconcentration and an increase in plasma osmolarity. Apparently, the excretion of water serves as a protective effect in relation to tissues that can be damaged against the background of its crystallization under the influence of cold.
The main stages of adaptation to cold
Long-term adaptation to cold has an ambiguous effect on the structural and functional rearrangements of the body. Along with the hypertrophy of the sympathetic-adrenal system, the thyroid gland, the mitochondrial system in the muscles and all links of oxygen transport, there is fatty malnutrition of the liver and a decrease in its detoxification functions, dystrophic phenomena from a number of systems with a decrease in their functional potential.
There are 4 stages of adaptation to cold
(N.A. Barbarash, G.Ya. Dvurechenskaya)
The first - emergency - unstable adaptation to cold
It is characterized by a sharp reaction of limiting heat transfer in the form of a spasm of peripheral vessels. The increase in heat production occurs due to the breakdown of ATP reserves and contractile thermogenesis. Deficiency of energy-rich phosphates develops. Damage may develop (frostbite, fermentemia, tissue necrosis).
The second - transitional - stage of urgent adaptation
There is a decrease in the stress response while maintaining the hyperfunction of the sympathetic-adrenal system and the thyroid gland. The processes of synthesis of nucleic acids and proteins, ATP resynthesis are activated. Vasoconstriction of peripheral tissues decreases, and, consequently, the risk of damage.
Third - sustainability - the stage of long-term adaptation
Long-term adaptation is formed with periodic exposure to cold. With its continuous exposure, it is less likely. It is characterized by hypertrophy of the sympathetic-adrenal system, thyroid gland, increased redox reactions, which leads to both direct adaptation to cold (stationary increase in heat production to maintain homeostasis), and positive cross - atherosclerosis, saline hypertension, hypoxia. Regulatory systems, including higher ones, become more resistant to stress.
Fourth stage - exhaustion
It develops with continuous prolonged or intense periodic exposure to cold. It is characterized by the phenomena of negative cross-adaptation, with the development of chronic diseases and dystrophic processes with a decrease in function in a number of internal organs.
Like any creature, the horse is able to adapt to the cold to some extent. The question is: how harmless to the health of the horse would such an adaptation be? What is the critical temperature? Are we sure that all horses react the same way to cold?
Even if we talk about a healthy horse, which is almost impossible after participating in sports or riding of any kind, is it as good in the cold, in rain and snow, as horse users of all faiths from athletes to naturists believe in it?
Thanks to "sports" veterinarians, we have a huge amount of research on the effect of heat and overheating on the horse - it's understandable: runs, races ... And there are too few serious works on the effect of cold on the body. Such studies can be counted on the fingers.
Here the trotters found out that at temperatures below -23 ° C, trotters die on the paths ... From cold air.
And when training in the cold at -22 ° C, they remain alive! From which it is concluded that at -22 ° C it is necessary to go out on the track, but in a blanket ...
For several years, the Finns found out in detail how Finnish horses freeze, measured the thickness of subcutaneous fat, the length of the hair - and found out that they are very cold. Conclusion: you need to wear blankets.
That's about all the research...
Of course, any attempt to study the effect of cold on the body will be incomplete until we know what the horse himself thinks about this.
In the meantime, there is no certainty that the horse actually feels in winter, we are forced to be guided by the strictly scientific data of anatomy and physiology and, of course, our own guesses and common sense. After all, our task is to make any weather of our not very gentle climate as comfortable as possible for horses.
Comfortable for a horse is considered to be a temperature from +24 to +5 ° C (in the absence of other irritating factors, of course). With this temperature regime, the horse does not need to expend additional energy on heating, provided that it is healthy and in good condition and in decent conditions.
Obviously, in any case, at temperatures below -GS, the horse will need additional sources of heat, and often, given the humidity, windiness, etc., such a need may arise even in the range of "comfortable" temperatures.
What is the body's physiological response to cold?
Immediate response. Occurs in response to a sudden sharp change in air temperature. The horse noticeably freezes, its hair stands on end (piloerection), the blood from the limbs drains to the internal organs - the legs, ears, nose become cold. The horse stands with its tail between its legs, not moving in order to save energy.
Adaptation. This is the next reaction of a horse exposed to further constant exposure to cold. It usually takes 10 to 21 days for a horse to get used to the cold. For example, a horse kept at a temperature of +20°C suddenly finds itself in conditions with a temperature of +5°C. It adapts to new environmental conditions in 21 days. With a further decrease in temperature from +5 to -5 ° C, the horse will need up to 21 days to adapt. And so on until the temperature reaches the lower critical point (LCR) of -15 ° C for an adult horse or 0 ° C for a growing one. Upon reaching a critical temperature, the horse's body will begin to work in an "emergency mode", not to live, but to survive, which will lead to a serious and, at times, irreversible, depletion of its resources.
As soon as the NCR is reached, stressful physiological changes begin, and the horse needs human intervention to cope with the cold: heating, additional nutrition.
It is clear that all data is conditional and differs for each specific horse. However, science does not yet have exact data.
Physiological changes consist in "focusing" the blood supply on the internal organs, the circulatory system begins to work, as it were, in a "small circle". There is a decrease in respiratory and cardiac rhythms to keep warm, resulting in a horse's inactivity in winter. The most visible outward sign of physiological change is the growth of a long, thick coat.
Fouling varies greatly in intensity from horse to horse under the same conditions. Breed, health, fatness, gender, type are of great importance. The more "thick-skinned" the horse, the heavier its type, the more it grows. As noted by N. D. Alekseev (1992), the Yakut horses have the thickest skin compared to horses of other breeds (4.4 + 0.05 mm in winter in the region of the last rib). Compare: in a European warm-blooded horse, the thickness of the skin in the same place is approximately 3-3.6 mm. There are exceptions related to the individual characteristics of metabolism. Temperament plays a role: active "thin-skinned" stallions of warm-blooded breeds are overgrown with little or no overgrowth at all. For example, Kao lives in the same conditions as our other horses, but does not overgrow at all - he walks in winter in summer wool. Ponies, heavy trucks, trotters, as a rule, grow stronger, they have pronounced “brushes”, hair growth from the wrist to the corolla increases significantly and a not very attractive, downright priestly beard appears. The same applies to sick and hungry horses - the body tries to compensate for the lack of a thermally insulating fat layer and malnutrition by spending the last reserves on growing hair, although here everything is strictly individual. By the length of a horse's coat, one can always accurately judge its health, maintenance and care.

In general, fouling seems to be a common thing for everyone ... But what does it cost a horse? I won’t say it better than my husband, so I’ll quote directly: “The process of fouling takes a significant part of the physiological forces. Just try to calculate what it costs the horse's body to raise, keep, tallow, etc. long hair. After all, it was not her husband who bought a fur coat for her, but she had to withdraw a very large "sum" from her own biological and physiological estate and spend it on wool, despite the fact that the biological resource of a horse is not so great. Nature has established a certain "warming standard" for a given strip (north, west, center of Russia). This standard can be easily calculated by analyzing the warming norms of wild animals that radically live in the natural environment of a given region, counting and analyzing the length of the coat, the depth and density of the undercoat, and the body temperature (normally) of these animals. This is a normal "natural" program that meets the requirements of the climate and the season. The man didn't interfere.
Through natural selection, this thermal standard and the standard of insulation has been developed for tens of thousands of years. It is precisely this amount of protective wool, precisely such density and depth of the undercoat, precisely such body temperature, as presented by the wild natural inhabitants of the region, that is the norm that ensures survival, and possibly some comfort.
The horse is not suitable here as a "trendsetter", being introduced, alien to this strip of being - no matter what generation. A kind of "lost exotic dog".
But for adaptive evolutionary changes, millennia are needed!
All that a horse can "present" to Russian cold weather is 2.5 - 3 cm of wool. No undercoat.
Having found out the discrepancy between the quality of horse insulation and local natural standards, we can speak with confidence about the physiological suffering of the horse, about causing both physiological and functional harm to the horse by cold. And this, and only this, will be a strictly scientific point of view. The argument based on the analysis of what is "worn in this band" for survival is irrefutable and very serious. Even two hours of a winter walk in the conditions of exposure to the natural climatic conditions of the North-West, unfortunately, is either very uncomfortable for the horse, or frankly dangerous.”
Send your good work in the knowledge base is simple. Use the form below
Students, graduate students, young scientists who use the knowledge base in their studies and work will be very grateful to you.
Hosted at http://www.allbest.ru/
Ministry of Sports and Tourism of the Republic of Belarus
educational institution
"Belarusian State University of Physical Culture"
Tourism Institute
Department of Technology in the Tourism Industry
Kontroll work
in the discipline "Physiology"
on thetopicsat" Adaptation to the action of low temperature"
Completed by: 2nd year student of 421 groups
part-time education
Faculty of Tourism and Hospitality
Tsinyavskaya Anastasia Viktorovna
Checked by: Bobr Vladimir Matveevich
- Introduction
- 1. Adaptation to low temperature
- 1.1 Physiological responses to exercise in low ambient temperatures
- 1.2 Metabolic reactions
- Conclusion
- List of used literature
Introduction
The human body is influenced by such a climatic factor as temperature. Temperature is one of the important abiotic factors affecting the physiological functions of all living organisms. The temperature depends on the geographic latitude, altitude, and season.
When temperature factors change, the human body produces specific adaptation reactions with respect to each factor. That is, it adapts.
Adaptation is a process of adaptation that is formed during a person's life. Thanks to adaptive processes, a person adapts to unusual conditions or a new level of activity, i.e. increases the resistance of his body against the action of various factors. The human body can adapt to high and low temperatures, low atmospheric pressure, or even some pathogenic factors.
People living in the northern or southern latitudes, in the mountains or on the plains, in the humid tropics or in the desert differ from each other in many indicators of homeostasis. Therefore, a number of normal indicators for individual regions of the globe may differ.
1. Adaptation to low temperature
Adaptation to cold - the most difficult - achievable and quickly lost without special training type of human climatic adaptation. This is explained by the fact that, according to modern scientific concepts, our ancestors lived in a warm climate and were much more adapted to protect themselves from overheating. The onset of cooling was relatively fast and man, as a species, "did not have time" to adapt to this climate change in most of the planet. In addition, people began to adapt to the conditions of low temperatures, mainly due to social and technogenic factors - housing, hearth, clothing. However, under extreme conditions of human activity (including climbing practice), the physiological mechanisms of thermoregulation - its "chemical" and "physical" sides become vital.
The first reaction of the body to the effects of cold is to reduce skin and respiratory (respiratory) heat loss due to vasoconstriction of the skin and pulmonary alveoli, as well as by reducing pulmonary ventilation (decrease in the depth and frequency of breathing). Due to changes in the lumen of the vessels of the skin, the blood flow in it can vary over a very wide range - from 20 ml to 3 liters per minute in the entire mass of the skin.
Vasoconstriction leads to a decrease in skin temperature, but when this temperature reaches 6°C and there is a threat of cold injury, the reverse mechanism develops - reactive hyperemia of the skin. With strong cooling, persistent vasoconstriction in the form of their spasm can occur. In this case, a signal of trouble appears - pain.
A decrease in the temperature of the skin of the hands to 27 °C is associated with a feeling of "cold", at a temperature below 20 °C - "very cold", at a temperature below 15 °C - "unbearably cold".
When exposed to cold, vasoconstructive (vasoconstrictive) reactions occur not only in cooled areas of the skin, but also in distant areas of the body, including internal organs ("reflected reaction"). Reflected reactions are especially pronounced when the feet are cooled - reactions of the nasal mucosa, respiratory organs, and internal genital organs. The vasoconstriction in this case causes a decrease in the temperature of the corresponding areas of the body and internal organs with the activation of the microbial flora. It is this mechanism that underlies the so-called "cold" diseases with the development of inflammation in the respiratory organs (pneumonia, bronchitis), urinary excretion (pyelitis, nephritis), genital area (adnexitis, prostatitis), etc.
The mechanisms of physical thermoregulation are the first to be included in the protection of the constancy of the internal environment when the balance of heat production and heat transfer is disturbed. If these reactions are not enough to maintain homeostasis, "chemical" mechanisms are activated - muscle tone increases, muscle tremors appear, which leads to an increase in oxygen consumption and an increase in heat production. At the same time, the work of the heart increases, blood pressure rises, and the speed of blood flow in the muscles increases. It has been calculated that in order to maintain the heat balance of a naked person with still cold air, it is necessary to increase heat production by 2 times for every 10º decrease in air temperature, and with a significant wind, heat production should double for every 5º decrease in air temperature. In a warmly dressed person, a doubling of the exchange value will compensate for a decrease in external temperature by 25º.
With repeated contacts with cold, local and general, a person develops protective mechanisms aimed at preventing the adverse effects of cold exposure. In the process of acclimatization to cold, resistance to frostbite increases (the frequency of frostbite in people acclimatized to cold is 6-7 times lower than in non-acclimatized people). In this case, first of all, there is an improvement in vasomotor mechanisms ("physical" thermoregulation). In persons exposed to cold for a long time, an increased activity of the processes of "chemical" thermoregulation is determined - the main metabolism; they have increased by 10 - 15%. Among the indigenous inhabitants of the North (for example, the Eskimos), this excess reaches 15 - 30% and is genetically fixed.
As a rule, in connection with the improvement of thermoregulation mechanisms in the process of acclimatization to cold, the share of participation of skeletal muscles in maintaining heat balance decreases - the intensity and duration of muscle trembling cycles become less pronounced. Calculations have shown that due to the physiological mechanisms of adaptation to cold, a naked person is able to endure air temperatures not lower than 2°C for a long time. Apparently, this air temperature is the limit of the body's compensatory capabilities to maintain the heat balance at a stable level.
The conditions under which the human body adapts to cold may be different (for example, work in unheated rooms, refrigeration units, outdoors in winter). At the same time, the effect of cold is not constant, but alternating with the temperature regime normal for the human body. Adaptation in such conditions is not clearly expressed. In the first days, reacting to low temperatures, heat generation increases uneconomically, heat transfer is still insufficiently limited. After adaptation, heat generation processes become more intense, and heat transfer decreases.
Otherwise, adaptation to the conditions of life in the northern latitudes occurs, where a person is affected not only by low temperatures, but also by the lighting regime and the level of solar radiation characteristic of these latitudes.
What happens in the human body during cooling?
As a result of irritation of cold receptors, the reflex reactions that regulate the preservation of heat change: the blood vessels of the skin narrow, which reduces the body's heat transfer by a third. It is important that the processes of heat generation and heat transfer are balanced. The predominance of heat transfer over heat generation leads to a decrease in body temperature and a violation of body functions. At a body temperature of 35 ° C, a mental disorder is observed. A further decrease in temperature slows down blood circulation, metabolism, and at temperatures below 25 ° C, breathing stops.
One of the factors of intensification of energy processes is lipid metabolism. For example, polar explorers, whose metabolism slows down in conditions of low air temperature, take into account the need to compensate for energy costs. Their diets are high in energy value (calorie content).
Residents of the northern regions have a more intense metabolism. The bulk of their diet is made up of proteins and fats. Therefore, in their blood, the content of fatty acids is increased, and the level of sugar is somewhat lowered.
People adapting to the humid, cold climate and oxygen deficiency of the North also have increased gas exchange, high cholesterol in the blood serum and mineralization of the bones of the skeleton, a thicker layer of subcutaneous fat (acting as a heat insulator).
However, not all people are equally adaptable. In particular, in some people in the conditions of the North, defense mechanisms and adaptive restructuring of the body can cause disadaptation - a whole series of pathological changes called "polar disease".
One of the most important factors that ensure human adaptation to the conditions of the Far North is the body's need for ascorbic acid (vitamin C), which increases the body's resistance to various infections.
The thermal insulation shell of our body includes the surface of the skin with subcutaneous fat, as well as the muscles located under it. When skin temperature falls below normal levels, constriction of the skin's blood vessels and contraction of skeletal muscles increase the insulating properties of the skin. It has been established that vasoconstriction of the passive muscle provides up to 85% of the total insulating capacity of the body under conditions of extremely low temperatures. This value of resistance to heat loss is 3-4 times higher than the insulating capacity of fat and skin.
1.1 Physiological responses to exercise in low ambient temperatures
metabolic temperature adaptation
As the muscle cools, it becomes weaker. The nervous system reacts to muscle cooling by changing the structure of the involvement of muscle fibers in the work. According to some experts, this change in the choice of fibers leads to a decrease in the efficiency of muscle contractions. At low temperatures, both the speed and strength of muscle contraction decrease. Attempting to perform work at a muscle temperature of 25°C with the same speed and productivity with which it was performed when the muscle temperature was 35°C will lead to rapid fatigue. Therefore, you have to either expend more energy or perform physical activity at a slower speed.
If clothing and exercise-induced metabolism are sufficient to maintain body temperature in cold environments, muscle performance will not decrease. However, as fatigue appears and muscle activity slows down, heat generation will gradually decrease.
1.2 Metabolic reactions
Prolonged exercise leads to increased utilization and oxidation of free fatty acids. Increased lipid metabolism is due mainly to the release of catecholamines (adrenaline and norepinephrine) into the vascular system. Under conditions of low ambient temperature, the secretion of these catecholamines is markedly increased, while the levels of free fatty acids rise much less compared to those during prolonged exercise in conditions of higher ambient temperature. Low ambient temperature causes constriction of blood vessels in the skin and subcutaneous tissues. As you know, the subcutaneous tissue is the main storage site for lipids (adipose tissue), so vasoconstriction leads to limited blood supply to the areas. From which free fatty acids are mobilized, so that the levels of free fatty acids do not rise as much.
Blood glucose plays an important role in the development of tolerance to low temperature conditions, as well as maintaining the level of endurance during exercise. loads. Hypoglycemia (low blood glucose), for example, suppresses shivering and leads to a significant drop in rectal temperature.
Many are interested in whether the airways are damaged by rapid deep inhalation of cold air. Cold air, passing through the mouth and trachea, quickly warms up, even if its temperature is below -25°C. Even at this temperature, the air, having passed about 5 cm along the nasal passage, warms up to 15 ° C. Very cold air, getting into the nose, warms up enough, approaching the exit from the nasal passage; thus, there is no danger of injury to the throat, trachea or lungs.
Conclusion
The conditions under which the body must adapt to the cold may be different. One of the possible options for such conditions is work in cold shops. In this case, the cold acts intermittently. In connection with the increased pace of development of the Far North, the issue of adapting the human body to life in the northern latitudes, where it is exposed not only to low temperatures, but also to changes in the illumination regime and radiation level, is currently becoming relevant.
Adaptive mechanisms make it possible to compensate for changes in the environmental factor only within certain limits and for a certain time. As a result of the impact on the body of factors that exceed the capabilities of adaptive mechanisms, maladjustment develops. It leads to dysfunction of the body systems. Consequently, there is a transition of an adaptive reaction into a pathological one - a disease. An example of diseases of maladjustment are cardiovascular diseases in non-native residents of the North.
List of used literature
1. Azhaev A.N., Berzin I.A., Deeva S.A., "Physiological and hygienic aspects of low temperatures on the human body", 2008
2. http://bibliofond.ru/view.aspx?id=459098#1
3. http://fiziologija.vse-zabolevaniya.ru/fiziologija-processov-adaptacii/ponjatie-adaptacii.html
4. http://human-physiology.ru/adaptaciya-ee-vidy-i-periody
Hosted on Allbest.ru
Similar Documents
The structure and functions of the skin. Basic mechanisms of thermoregulation. Skin response to ambient temperature. Is the body always able to compensate for prolonged exposure to low or high temperature. First aid for heat and sunstroke.
presentation, added 12/02/2013
The main causes of death of plants from the cold. Instantaneous and irreversible damage to cells during the formation of intracellular ice as an indication of the physical nature of the process. Exposure of membranes to hypothermia, ways to prevent it.
abstract, added 08/11/2009
Adaptation as one of the key concepts in human ecology. The main mechanisms of human adaptation. Physiological and biochemical bases of adaptation. Adaptation of the body to physical stress. Decreased excitability during the development of limiting inhibition.
abstract, added 06/25/2011
Characterization of the processes of human adaptation to environmental conditions. Study of the main mechanisms of adaptation. The study of general measures to increase the resistance of the body. Laws and patterns of hygiene. Descriptions of the principles of hygienic regulation.
presentation, added 03/11/2014
The study of the concept of physical and chemical thermoregulation. Isothermia - the constancy of body temperature. Factors affecting body temperature. Causes and signs of hypothermia and hyperthermia. Temperature measuring points. Types of fevers. Hardening of the body.
presentation, added 10/21/2013
Features of the habitat of amphibians (frogs, toads, newts and salamanders). Dependence of the body temperature of amphibians on the ambient temperature. The benefits of amphibians for agriculture. Orders of amphibians: legless, tailless and tailed.
presentation, added 02/28/2011
The importance of maintaining a constant temperature of the internal environment of the body (isothermia) to ensure vital processes. Physical thermoregulation, which occurs by changing the release of heat from the body. The role of hormones in chemical thermoregulation.
presentation, added 04/18/2019
Cross-adaptation of an organism to one environmental factor, its promotion of adaptation to other factors. Molecular basis of human adaptation and its practical significance. The adaptability of a living organism to damaging environmental factors.
abstract, added 20.09.2009
Adaptation of an organism to environmental conditions in general biological terms, its necessity for the preservation of both the individual and the species. Methods of protection from adverse environmental conditions. Anabiosis, stupor, hibernation, migration, enzyme activation.
abstract, added 20.09.2009
Adaptation is the adaptation of an organism to its environment, to the conditions of its existence. Features of the living conditions of an athlete. Biochemical and physiological mechanisms of adaptation to physical stress. Biological principles of sports training.