With my vision of how the brain works and what are the possible ways to create artificial intelligence. Since then, significant progress has been made. Some things were better understood, some things were modeled on a computer. What’s nice is that there are like-minded people actively participating in the project.
In this series of articles we plan to talk about the concept of intelligence that we are currently working on and demonstrate some solutions that are fundamentally new in the field of modeling the functioning of the brain. But in order for the narrative to be clear and consistent, it will contain not only a description of new ideas, but also a story about the work of the brain in general. Some things, especially at the beginning, may seem simple and well-known, but I would advise you not to skip them, since they largely determine the overall evidence of the narrative.
Understanding the Brain
Nerve cells, also known as neurons, together with their fibers that transmit signals, form the nervous system. In vertebrates, the bulk of neurons are concentrated in the cranial cavity and spinal canal. This is called the central nervous system. Accordingly, the brain and spinal cord are distinguished as its components.
The spinal cord collects signals from most of the body's receptors and transmits them to the brain. Through the structures of the thalamus, they are distributed and projected onto the cerebral cortex.
In addition to the cerebral hemispheres, the cerebellum, which is essentially a small independent brain, also processes information. The cerebellum provides fine motor skills and coordination of all movements.
Vision, hearing and smell provide the brain with a stream of information about the outside world. Each of the components of this flow, having passed along its own tract, is also projected onto the cortex. The cortex is a layer of gray matter, 1.3 to 4.5 mm thick, that makes up the outer surface of the brain. Due to the convolutions formed by folds, the bark is packed so that it occupies three times less area than when flattened. The total area of the cortex of one hemisphere is approximately 7000 sq.cm.
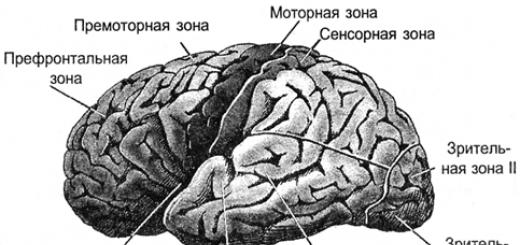
As a result, all signals are projected onto the cortex. The projection is carried out by bundles of nerve fibers that are distributed over limited areas of the cortex. The area onto which either external information or information from other parts of the brain is projected forms the cortex zone. Depending on what signals such a zone receives, it has its own specialization. There are motor cortex zones, sensory zones, Broca's and Wernicke's areas, visual zones, occipital lobe, about a hundred different zones in total.

In the vertical direction, the bark is usually divided into six layers. These layers do not have clear boundaries and are determined by the predominance of one or another cell type. In different zones of the cortex, these layers can be expressed differently, stronger or weaker. But, in general, we can say that the cortex is quite universal, and assume that the functioning of its different zones obeys the same principles.

Layers of bark
Signals travel through afferent fibers to the cortex. They reach the III and IV levels of the cortex, where they are distributed among the neurons closest to the place where the afferent fiber entered. Most neurons have axon connections within their cortical area. But some neurons have axons that extend beyond it. Along these efferent fibers, signals go either outside the brain, for example, to the executive organs, or are projected to other parts of the cortex of their own or another hemisphere. Depending on the direction of signal transmission, efferent fibers are usually divided into:
- associative fibers that connect individual areas of the cortex of one hemisphere;
- commissural fibers that connect the cortex of the two hemispheres;
- projection fibers that connect the cortex with the nuclei of the lower parts of the central nervous system.
You can imagine the cerebral cortex as a large canvas, cut into separate zones. The pattern of neuron activity in each zone encodes certain information. Bundles of nerve fibers formed by axons extending beyond their cortical zone form a system of projection connections. Certain information is projected onto each zone. Moreover, one zone can simultaneously receive several information flows, which can come from both zones of its own and the opposite hemisphere. Each stream of information is like a unique picture drawn by the activity of the axons of the nerve bundle. The functioning of a separate zone of the cortex is the receipt of multiple projections, memorization of information, its processing, the formation of its own picture of activity and the further projection of information resulting from the work of this zone.
A significant amount of the brain is white matter. It is formed by the axons of neurons, creating those same projection paths. In the picture below, white matter can be seen as a light fill between the cortex and the internal structures of the brain.

Distribution of white matter in a frontal section of the brain
Using diffuse spectral MRI, it was possible to track the direction of individual fibers and build a three-dimensional model of the connectivity of cortical zones (Connectomics project).
The figures below give a good idea of the structure of the connections (Van J. Wedeen, Douglas L. Rosene, Ruopeng Wang, Guangping Dai, Farzad Mortazavi, Patric Hagmann, Jon H. Kaas, Wen-Yih I. Tseng, 2012).

View from the left hemisphere

Back view

Right view
By the way, in the rear view, the asymmetry of the projection paths of the left and right hemispheres is clearly visible. This asymmetry largely determines the differences in the functions that the hemispheres acquire as they learn.
Neuron
The basis of the brain is the neuron. Naturally, modeling the brain using neural networks begins with answering the question of what is the principle of its operation.
The operation of a real neuron is based on chemical processes. At rest, there is a potential difference between the internal and external environment of the neuron - the membrane potential, which is about 75 millivolts. It is formed due to the work of special protein molecules that act as sodium-potassium pumps. These pumps, using the energy of the ATP nucleotide, drive potassium ions inside and sodium ions out of the cell. Since the protein acts as an ATPase, that is, an enzyme that hydrolyzes ATP, it is called “sodium-potassium ATPase.” As a result, the neuron turns into a charged capacitor with a negative charge on the inside and a positive charge on the outside.

Neuron diagram (Mariana Ruiz Villarreal)
The surface of a neuron is covered with branching processes called dendrites. The dendrites are adjacent to the axon endings of other neurons. The places where they connect are called synapses. Through synaptic interaction, a neuron is able to respond to incoming signals and, under certain circumstances, generate its own impulse, called a spike.
Signal transmission at synapses occurs due to substances called neurotransmitters. When a nerve impulse enters a synapse along an axon, it releases neurotransmitter molecules characteristic of this synapse from special vesicles. On the membrane of the neuron receiving the signal there are protein molecules - receptors. Receptors interact with neurotransmitters.

Chemical synapse
Receptors located in the synaptic cleft are ionotropic. This name emphasizes the fact that they are also ion channels capable of moving ions. Neurotransmitters act on receptors in such a way that their ion channels open. Accordingly, the membrane either depolarizes or hyperpolarizes, depending on which channels are affected and, accordingly, what type of synapse it is. In excitatory synapses, channels open that allow cations to enter the cell - the membrane is depolarized. In inhibitory synapses, channels that conduct anions open, which leads to hyperpolarization of the membrane.
Under certain circumstances, synapses can change their sensitivity, which is called synaptic plasticity. This leads to the fact that the synapses of one neuron acquire different susceptibility to external signals.
At the same time, many signals arrive at the synapses of a neuron. Inhibitory synapses pull the membrane potential towards the accumulation of charge inside the cell. Activating synapses, on the contrary, try to discharge the neuron (figure below).

Excitation (A) and inhibition (B) of the retinal ganglion cell (Nicholls J., Martin R., Wallas B., Fuchs P., 2003)
When the total activity exceeds the initiation threshold, a discharge called an action potential or spike occurs. A spike is a sharp depolarization of the neuron membrane, which generates an electrical impulse. The entire pulse generation process lasts about 1 millisecond. Moreover, neither the duration nor the amplitude of the impulse depends on how strong the reasons that caused it were (figure below).

Recording the action potential of a ganglion cell (Nicholls J., Martin R., Wallas B., Fuchs P., 2003)
After the spike, ion pumps ensure reuptake of the neurotransmitter and clearing of the synaptic cleft. During the refractory period that occurs after a spike, the neuron is not able to generate new impulses. The duration of this period determines the maximum firing frequency that the neuron is capable of.
Spikes that occur as a consequence of activity at synapses are called evoked. The evoked spike rate encodes how well the incoming signal matches the sensitivity setting of the neuron's synapses. When incoming signals arrive precisely at the sensitive synapses that activate the neuron, and this is not interfered with by signals arriving at the inhibitory synapses, the neuron’s response is maximum. The image that is described by such signals is called a characteristic stimulus for the neuron.
Of course, the idea of how neurons work should not be oversimplified. Information between some neurons can be transmitted not only by spikes, but also by channels connecting their intracellular contents and transmitting electrical potential directly. This propagation is called gradual, and the connection itself is called an electrical synapse. Dendrites, depending on the distance to the neuron body, are divided into proximal (close) and distal (remote). Distal dendrites can form sections that act as semi-autonomous elements. In addition to synaptic excitation pathways, there are extrasynaptic mechanisms that cause metabotropic spikes. In addition to evoked activity, there is also spontaneous activity. Finally, brain neurons are surrounded by glial cells, which also have a significant influence on ongoing processes.
The long path of evolution has created many mechanisms that the brain uses in its work. Some of them can be understood on their own; the meaning of others becomes clear only when considering rather complex interactions. Therefore, you should not take the above description of the neuron as exhaustive. To move on to deeper models, we need to first understand the “basic” properties of neurons.
In 1952, Alan Lloyd Hodgkin and Andrew Huxley described the electrical mechanisms that determine the generation and transmission of nerve signals in the squid giant axon (Hodgkin, 1952). Which was awarded the Nobel Prize in Physiology or Medicine in 1963. The Hodgkin–Huxley model describes the behavior of a neuron using a system of ordinary differential equations. These equations correspond to the autowave process in the active medium. They take into account many components, each of which has its own biophysical analogue in a real cell (figure below). Ion pumps correspond to the current source I p . The inner lipid layer of the cell membrane forms a capacitor with capacitance Cm. The ion channels of synaptic receptors provide electrical conductivity g n, which depends on the signals supplied, varying with time t, and the overall value of the membrane potential V. The leakage current of the membrane pores creates a conductor g L. The movement of ions through ion channels occurs under the influence of electrochemical gradients, which correspond to voltage sources with electromotive forces E n and E L .

Main components of the Hodgkin-Huxley model
Naturally, when creating neural networks, there is a desire to simplify the neuron model, leaving only the most essential properties in it. The most famous and popular simplified model is the McCulloch-Pitts artificial neuron, developed in the early 1940s (McCulloch J., Pitts W., 1956). 
Formal McCulloch-Pitts neuron
Signals are sent to the inputs of such a neuron. These signals are weighted summed. Next, some nonlinear activation function, for example, sigmoidal, is applied to this linear combination. The logistic function is often used as a sigmoid function: ![]()

Logistic function
In this case, the activity of a formal neuron is written as

As a result, such a neuron turns into a threshold adder. With a sufficiently steep threshold function, the neuron's output signal is either 0 or 1. The weighted sum of the input signal and the neuron's weights is the convolution of two images: the image of the input signal and the image described by the neuron's weights. The more accurate the match between these images, the higher the convolution result. That is, the neuron essentially determines how similar the supplied signal is to the image recorded at its synapses. When the convolution value exceeds a certain level and the threshold function switches to one, this can be interpreted as a decisive statement by the neuron that it has recognized the presented image.
Real neurons are indeed somewhat similar to McCulloch-Pitts neurons. The amplitude of their spikes does not depend on what signals at the synapses caused them. Spike either exists or he doesn't. But real neurons respond to a stimulus not with a single impulse, but with a sequence of impulses. In this case, the frequency of impulses is higher, the more accurately the characteristic image of a neuron is recognized. This means that if we build a neural network from such threshold adders, then, with a static input signal, although it will give some output result, this result will be far from reproducing how real neurons work. In order to bring a neural network closer to a biological prototype, we will need to simulate the work in dynamics, taking into account time parameters and reproducing the frequency properties of signals.
But you can go another way. For example, we can identify a generalized characteristic of the activity of a neuron, which corresponds to the frequency of its impulses, that is, the number of spikes over a certain period of time. If we go to this description, we can imagine a neuron as a simple linear adder.

Linear adder
The output and, accordingly, input signals for such neurons are no longer dichatomic (0 or 1), but are expressed by a certain scalar quantity. The activation function is then written as
A linear adder should not be perceived as something fundamentally different compared to a spiking neuron; it simply allows one to move to longer time intervals when modeling or describing them. And although the pulse description is more correct, the transition to a linear adder in many cases is justified by a strong simplification of the model. Moreover, some important properties that are difficult to see in a spiking neuron are quite obvious in a linear adder.
This cell has a complex structure, is highly specialized and in structure contains a nucleus, a cell body and processes. There are more than one hundred billion neurons in the human body.
Review
The complexity and variety of functions of the nervous system are determined by the interactions between neurons, which, in turn, represent a set of different signals transmitted as part of the interaction of neurons with other neurons or muscles and glands. Signals are emitted and propagated by ions that generate an electrical charge that travels along the neuron.
Structure
A neuron consists of a body with a diameter of 3 to 130 µm, containing a nucleus (with a large number of nuclear pores) and organelles (including a highly developed rough ER with active ribosomes, the Golgi apparatus), as well as processes. There are two types of processes: dendrites and . The neuron has a developed and complex cytoskeleton that penetrates its processes. The cytoskeleton maintains the shape of the cell; its threads serve as “rails” for the transport of organelles and substances packaged in membrane vesicles (for example, neurotransmitters). The cytoskeleton of a neuron consists of fibrils of different diameters: Microtubules (D = 20-30 nm) - consist of the protein tubulin and stretch from the neuron along the axon, right up to the nerve endings. Neurofilaments (D = 10 nm) - together with microtubules provide intracellular transport of substances. Microfilaments (D = 5 nm) - consist of the proteins actin and myosin, especially pronounced in growing nerve processes and in. A developed synthetic apparatus is revealed in the body of the neuron; the granular ER of the neuron is stained basophilically and is known as the “tigroid”. The tigroid penetrates the initial sections of the dendrites, but is located at a noticeable distance from the beginning of the axon, which serves as a histological sign of the axon.
There is a distinction between anterograde (away from the body) and retrograde (toward the body) axon transport.
Dendrites and axon
An axon is usually a long process adapted to conduct from the body of a neuron. Dendrites are, as a rule, short and highly branched processes that serve as the main site of formation of excitatory and inhibitory synapses influencing the neuron (different neurons have different ratios of axon and dendrites lengths). A neuron may have several dendrites and usually only one axon. One neuron can have connections with many (up to 20 thousand) other neurons.
Dendrites divide dichotomously, while axons give off collaterals. Mitochondria are usually concentrated at branching nodes.
Dendrites do not have a myelin sheath, but axons may have one. The place of generation of excitation in most neurons is the axon hillock - a formation at the point where the axon departs from the body. In all neurons, this zone is called the trigger zone.
Synapse(Greek σύναψις, from συνάπτειν - hug, clasp, shake hands) - the place of contact between two neurons or between a neuron and the effector cell receiving the signal. Serves for transmission between two cells, and during synaptic transmission the amplitude and frequency of the signal can be adjusted. Some synapses cause depolarization of the neuron, others hyperpolarization; the former are excitatory, the latter are inhibitory. Typically, stimulation from several excitatory synapses is necessary to excite a neuron.
The term was introduced in 1897 by the English physiologist Charles Sherrington.
Classification
Structural classification
Based on the number and arrangement of dendrites and axons, neurons are divided into axonless neurons, unipolar neurons, pseudounipolar neurons, bipolar neurons, and multipolar (many dendritic arbors, usually efferent) neurons.
Axonless neurons- small cells, grouped nearby in the intervertebral ganglia, without anatomical signs of division of processes into dendrites and axons. All processes of the cell are very similar. The functional purpose of axonless neurons is poorly understood.
Unipolar neurons- neurons with one process, present, for example, in the sensory nucleus of the trigeminal nerve in.
Bipolar neurons- neurons having one axon and one dendrite, located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia.
Multipolar neurons- neurons with one axon and several dendrites. This type of nerve cells predominates in.
Pseudounipolar neurons- are unique in their kind. One process extends from the body, which immediately divides in a T-shape. This entire single tract is covered with a myelin sheath and is structurally an axon, although along one of the branches the excitation goes not from, but to the body of the neuron. Structurally, dendrites are branches at the end of this (peripheral) process. The trigger zone is the beginning of this branching (that is, it is located outside the cell body). Such neurons are found in the spinal ganglia.
Functional classification
Based on their position in the reflex arc, afferent neurons (sensitive neurons), efferent neurons (some of them are called motor neurons, sometimes this not very accurate name applies to the entire group of efferents) and interneurons (interneurons) are distinguished.
Afferent neurons(sensitive, sensory or receptor). Neurons of this type include primary cells and pseudounipolar cells, whose dendrites have free endings.
Efferent neurons(effector, motor or motor). Neurons of this type include the final neurons - ultimatum and penultimate - non-ultimatum.
Association neurons(intercalary or interneurons) - a group of neurons communicates between efferent and afferent ones; they are divided into intrusive, commissural and projection.
Secretory neurons- neurons that secrete highly active substances (neurohormones). They have a well-developed Golgi complex, the axon ends at axovasal synapses.
Morphological classification
The morphological structure of neurons is diverse. In this regard, several principles are used when classifying neurons:
- take into account the size and shape of the neuron body;
- number and nature of branching of processes;
- the length of the neuron and the presence of specialized membranes.
According to the shape of the cell, neurons can be spherical, granular, stellate, pyramidal, pear-shaped, fusiform, irregular, etc. The size of the neuron body varies from 5 μm in small granular cells to 120-150 μm in giant pyramidal neurons. The length of a human neuron ranges from 150 µm to 120 cm.
Based on the number of processes, the following morphological types of neurons are distinguished:
- unipolar (with one process) neurocytes, present, for example, in the sensory nucleus of the trigeminal nerve in;
- pseudounipolar cells grouped nearby in the intervertebral ganglia;
- bipolar neurons (have one axon and one dendrite), located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia;
- multipolar neurons (have one axon and several dendrites), predominant in the central nervous system.
Neuron development and growth
A neuron develops from a small precursor cell that stops dividing even before it produces its processes. (However, the issue of neuronal division currently remains controversial) As a rule, the axon begins to grow first, and dendrites form later. At the end of the developing process of the nerve cell, an irregularly shaped thickening appears, which, apparently, makes its way through the surrounding tissue. This thickening is called the growth cone of the nerve cell. It consists of a flattened part of the nerve cell process with many thin spines. The microspinuses are 0.1 to 0.2 µm thick and can reach 50 µm in length; the wide and flat region of the growth cone is about 5 µm in width and length, although its shape can vary. The spaces between the microspines of the growth cone are covered with a folded membrane. Microspines are in constant motion - some are retracted into the growth cone, others elongate, deviate in different directions, touch the substrate and can stick to it.
The growth cone is filled with small, sometimes connected to each other, membrane vesicles of irregular shape. Directly below the folded areas of the membrane and in the spines is a dense mass of entangled actin filaments. The growth cone also contains mitochondria, microtubules and neurofilaments found in the body of the neuron.
It is likely that microtubules and neurofilaments elongate mainly due to the addition of newly synthesized subunits at the base of the neuron process. They move at a speed of about a millimeter per day, which corresponds to the speed of slow axonal transport in a mature neuron. Since the average speed of advancement of the growth cone is approximately the same, it is possible that during the growth of the neuron process, neither the assembly nor destruction of microtubules and neurofilaments occurs at its far end. New membrane material is added, apparently, at the end. The growth cone is an area of rapid exocytosis and endocytosis, as evidenced by the many vesicles present there. Small membrane vesicles are transported along the neuron process from the cell body to the growth cone with a stream of fast axonal transport. The membrane material is apparently synthesized in the body of the neuron, transported to the growth cone in the form of vesicles and incorporated here into the plasma membrane by exocytosis, thus lengthening the process of the nerve cell.
The growth of axons and dendrites is usually preceded by a phase of neuronal migration, when immature neurons disperse and find a permanent home.
The structural unit of the nervous system is the nerve cell, or neuron. Neurons differ from other cells in the body in many ways. First of all, their population, numbering from 10 to 30 billion (and perhaps more*) cells, is almost completely “complete” by the time of birth, and not a single neuron, if it dies, is replaced by a new one. It is generally accepted that after a person passes the period of maturity, about 10 thousand neurons die every day, and after 40 years this daily figure doubles.
* The assumption that the nervous system consists of 30 billion neurons was made by Powell and his colleagues (Powell et al., 1980), who showed that in mammals, regardless of species, there are about 146 thousand nerve cells per 1 mm 2 of nervous tissue. The total surface area of the human brain is 22 dm 2 (Changeux, 1983, p. 72).
Another feature of neurons is that, unlike other types of cells, they do not produce, secrete or structure anything; their only function is to conduct neural information.
Neuron structure
There are many types of neurons, the structure of which varies depending on the functions they perform in the nervous system; a sensory neuron differs in structure from a motor neuron or a neuron of the cerebral cortex (Fig. A.28).
Rice. A.28. Different types of neurons.
But whatever the function of a neuron, all neurons are made up of three main parts: the cell body, dendrites and axon.
Body neuron, Like any other cell, it consists of cytoplasm and nucleus. The cytoplasm of a neuron, however, is especially rich mitochondria, responsible for producing the energy necessary to maintain high cell activity. As already noted, clusters of neuron bodies form nerve centers in the form of a ganglion, in which the number of cell bodies is in the thousands, a nucleus, where there are even more of them, or, finally, a cortex consisting of billions of neurons. The cell bodies of neurons form the so-called Gray matter.
Dendrites serve as a kind of antenna for the neuron. Some neurons have many hundreds of dendrites that receive information from receptors or other neurons and conduct it to the cell body and its only other type of process. - axon.
Axon is the part of a neuron responsible for transmitting information to the dendrites of other neurons, muscles or glands. In some neurons, the axon length reaches a meter, in others the axon is very short. As a rule, the axon branches, forming the so-called terminal tree; at the end of each branch there is synoptic plaque. It is she who forms the connection (synapse) of a given neuron with the dendrites or cell bodies of other neurons.
Most nerve fibers (axons) are covered with a sheath consisting of myelin- a white fat-like substance that acts as an insulating material. The myelin sheath is interrupted by constrictions at regular intervals of 1-2 mm - interceptions of Ranvier, which increase the speed of a nerve impulse traveling along a fiber, allowing it to “jump” from one interception to another, rather than gradually spreading along the fiber. Hundreds and thousands of axons collected in bundles form nerve pathways, which, thanks to myelin, have the appearance white matter.
Nerve impulse
Information enters the nerve centers, is processed there and then transmitted to effectors in the form nerve impulses, running along neurons and the nerve pathways connecting them.
Regardless of what information is transmitted by nerve impulses running along billions of nerve fibers, they are no different from each other. Why, then, do impulses coming from the ear convey information about sounds, and impulses from the eye convey information about the shape or color of an object, and not about sounds or something completely different? Yes, simply because the qualitative differences between nerve signals are determined not by these signals themselves, but by the place where they arrive: if it is a muscle, it will contract or stretch; if it is a gland, it will secrete, reduce or stop secretion; if this is a certain area of the brain, a visual image of an external stimulus will be formed in it, or the signal will be deciphered in the form of, for example, sounds. Theoretically, it would be enough to change the course of nerve pathways, for example, part of the optic nerve to the area of the brain responsible for deciphering sound signals, to force the body to “hear with the eyes.”
Resting potential and action potential
Nerve impulses are transmitted along dendrites and axons not by the external stimulus itself or even its energy. An external stimulus only activates the corresponding receptors, and this activation is converted into energy electric potential, which is created at the tips of dendrites that form contacts with the receptor.
The nerve impulse that arises can be roughly compared to fire running along a fuse and igniting a dynamite cartridge located in its path; The “fire” thus spreads towards the final target through small, successive explosions. The transmission of a nerve impulse, however, is fundamentally different from this in that almost immediately after the passage of the discharge, the potential of the nerve fiber is restored.
A nerve fiber at rest can be likened to a small battery; on the outside of its membrane there is a positive charge, and on the inside there is a negative charge (Fig. A.29), and this resting potential is converted into electric current only when both poles are closed. This is exactly what happens during the passage of a nerve impulse, when the fiber membrane for a moment becomes permeable and depolarized. Following this depolarization the period is coming refractoriness, during which the membrane repolarizes and restores the ability to conduct a new impulse*. So, due to successive depolarizations, this propagation occurs action potential(i.e., nerve impulse) at a constant speed, varying from 0.5 to 120 meters per second, depending on the type of fiber, its thickness and the presence or absence of a myelin sheath.
* During the refractory period, which lasts about a thousandth of a second, nerve impulses cannot travel along the fiber. Therefore, in one second, a nerve fiber is capable of conducting no more than 1000 impulses.

Rice. A.29. Action potential. The development of the action potential, accompanied by a change in electrical voltage (from -70 to + 40 mV), is due to the restoration of equilibrium between positive and negative ions on both sides of the membrane, the permeability of which increases for a short time.
The law "everything" or nothing". Since each nerve fiber has a certain electrical potential, the impulses propagating along it, regardless of the intensity or any other properties of the external stimulus, always have the same characteristics. This means that an impulse in a neuron can only occur if its activation, caused by stimulation of a receptor or an impulse from another neuron, exceeds a certain threshold below which activation is ineffective; but, if the threshold is reached, a “full” impulse immediately arises. This fact is called the “all or nothing” law.
Synaptic transmission
Synapse. A synapse is the area of connection between the axon terminal of one neuron and the dendrites or body of another. Each neuron can form up to 800-1000 synapses with other nerve cells, and the density of these contacts in the gray matter of the brain is more than 600 million per 1 mm 3 (Fig. A.30)*.
*This means that if you count 1000 synapses in one second, then it will take from 3 to 30 thousand years to completely recount them (Changeux, 1983, p. 75).

Rice. A.30. Synaptic connection of neurons (in the middle - the synapse area at higher magnification). The terminal plaque of the presynaptic neuron contains vesicles with a supply of neurotransmitter and mitochondria that supply the energy necessary for transmission of the nerve signal.
The place where a nerve impulse passes from one neuron to another is, in fact, not a point of contact, but rather a narrow gap called synoptic gap. We are talking about a gap with a width of 20 to 50 nanometers (millionths of a millimeter), which is limited on one side by the membrane of the presynaptic plaque of the neuron transmitting the impulse, and on the other by the postsynaptic membrane of the dendrite or body of another neuron, which receives the nerve signal and then transmits it further.
Neurotransmitters. It is at synapses that processes occur as a result of which chemicals released by the presynaptic membrane transmit a nerve signal from one neuron to another. These substances, called neurotransmitters(or simply mediators), a kind of “brain hormones” (neurohormones), accumulate in the vesicles of synaptic plaques and are released when a nerve impulse arrives here along the axon.
After this, the mediators diffuse into the synaptic cleft and attach to specific receptor sites postsynaptic membrane, i.e. to such areas to which they “fit like a key to a lock.” As a result, the permeability of the postsynaptic membrane changes, and thus the signal is transmitted from one neuron to another; Mediators can also block the transmission of nerve signals at the synapse level, reducing the excitability of the postsynaptic neuron.
Having fulfilled their function, the mediators are broken down or neutralized by enzymes or absorbed back into the presynaptic ending, which leads to the restoration of their supply in the vesicles by the time the next impulse arrives (Fig. A.31).

Rice. A.31. la. Mediator A, whose molecules are released from the terminal plaque of neuron I, binds to specific receptors on the dendrites of neuron II. X molecules, which in their configuration do not fit these receptors, cannot occupy them and therefore do not cause any synaptic effects.
1b. M molecules (for example, the molecules of some psychotropic drugs) are similar in configuration to molecules of the neurotransmitter A and therefore can bind to receptors for this neurotransmitter, thus preventing it from performing its functions. For example, LSD interferes with serotonin's ability to suppress sensory signals.
2a and 2b. Certain substances, called neuromodulators, can act at the axon terminal to facilitate or inhibit neurotransmitter release.
The excitatory or inhibitory function of a synapse depends mainly on the type of transmitter it secretes and on the effect of the latter on the postsynaptic membrane. Some mediators always have only an excitatory effect, others only have an inhibitory effect, and still others play the role of activators in some parts of the nervous system, and inhibitors in others.
Main functions neurotransmitters. Currently, several dozen of these neurohormones are known, but their functions have not yet been sufficiently studied. This applies, for example, to acetylcholine, which is involved in muscle contraction, causes a slowdown in heart and respiratory rates and is inactivated by an enzyme acetylcholinesterase*. The functions of such substances from the group are not fully understood monoamines, as norepinephrine, which is responsible for the wakefulness of the cerebral cortex and increased heart rate, dopamine, present in the "pleasure centers" of the limbic system and some nuclei of the reticular formation, where it participates in the processes of selective attention, or serotonin, which regulates sleep and determines the amount of information circulating in the sensory pathways. Partial inactivation of monoamines occurs as a result of their oxidation by the enzyme monoamine oxidase. This process, which usually returns brain activity to a normal level, in some cases can lead to an excessive decrease in it, which in psychological terms manifests itself in a person in a feeling of depression (depression).
* Apparently, a lack of acetylcholine in some nuclei of the diencephalon is one of the main causes of Alzheimer's disease, and a lack of dopamine in the putamen (one of the basal ganglia) may be the cause of Parkison's disease.
Gamma-aminobutyric acid (GABA) is a neurotransmitter that performs approximately the same physiological function as monoamine oxidase. Its action consists mainly of reducing the excitability of brain neurons in relation to nerve impulses.
Along with neurotransmitters, there is a group of so-called neuromodulators, which are mainly involved in the regulation of the nervous response, interacting with neurotransmitters and modifying their effects. As an example we can name substance P And bradykinin, involved in the transmission of pain signals. The release of these substances at spinal cord synapses, however, can be suppressed by secretion endorphins And enkephalin, which thus leads to a decrease in the flow of pain nerve impulses (Fig. A.31, 2a). The functions of modulators are also performed by substances such as factorS, apparently playing an important role in sleep processes, cholecystokinin, responsible for the feeling of satiety, angiotensin, thirst regulating, and other agents.
Neurotransmitters and the effect of psychotropic substances. It is now known that various psychotropic drugs act at the level of synapses and those processes in which neurotransmitters and neuromodulators participate.
The molecules of these drugs are similar in structure to the molecules of certain mediators, which allows them to “deceive” various mechanisms of synaptic transmission. Thus, they disrupt the action of true neurotransmitters, either taking their place at the receptor sites, or preventing them from being absorbed back into the presynaptic endings or being destroyed by specific enzymes (Fig. A.31, 26).
It has been established, for example, that LSD, by occupying serotonin receptor sites, prevents serotonin from inhibiting the influx of sensory signals. In this way, LSD opens the mind to a wide variety of stimuli that continually assault the senses.
Cocaine enhances the effects of dopamine, taking its place in receptor sites. They act in a similar way morphine and other opiates, the immediate effect of which is explained by the fact that they quickly manage to occupy receptor sites for endorphins*.
* Accidents associated with drug overdose are explained by the fact that the binding of excessive amounts of, for example, heroin by zndorphin receptors in the nerve centers of the medulla oblongata leads to a sharp depression of breathing, and sometimes to a complete stop (Besson, 1988, Science et Vie, Hors serie, n° 162).
Action amphetamines due to the fact that they suppress the reuptake of norepinephrine by presynaptic endings. As a result, the accumulation of excess amounts of neurohormone in the synaptic cleft leads to an excessive degree of wakefulness in the cerebral cortex.
It is generally accepted that the effects of the so-called tranquilizers(for example, Valium) are explained mainly by their facilitating effect on the action of GABA in the limbic system, which leads to increased inhibitory effects of this neurotransmitter. On the contrary, how antidepressants These are mainly enzymes that inactivate GABA, or drugs such as, for example, monoamine oxidase inhibitors, the introduction of which increases the amount of monoamines in synapses.
Death by some poisonous gases occurs due to suffocation. This effect of these gases is due to the fact that their molecules block the secretion of an enzyme that destroys acetylcholine. Meanwhile, acetylcholine causes muscle contraction and a slowdown in heart and respiratory rates. Therefore, its accumulation in synaptic spaces leads to inhibition and then complete blockade of cardiac and respiratory functions and a simultaneous increase in the tone of all muscles.
The study of neurotransmitters is just beginning, and we can expect that hundreds, and perhaps thousands of these substances will soon be discovered, the diverse functions of which determine their primary role in the regulation of behavior.
An axon is usually a long extension of a neuron, adapted to conduct excitation and information from the body of the neuron or from the neuron to the executive organ. Dendrites are, as a rule, short and highly branched processes of a neuron, serving as the main site of formation of excitatory and inhibitory synapses affecting the neuron (different neurons have different ratios of axon and dendrites lengths), and which transmit excitation to the neuron body. A neuron may have several dendrites and usually only one axon. One neuron can have connections with many (up to 20 thousand) other neurons.
Dendrites divide dichotomously, while axons give off collaterals. Mitochondria are usually concentrated at branching nodes.
Dendrites do not have a myelin sheath, but axons may have one. The place of generation of excitation in most neurons is the axon hillock - a formation at the point where the axon departs from the body. In all neurons, this zone is called the trigger zone.
A synapse is a point of contact between two neurons or between a neuron and an effector cell receiving a signal. It serves to transmit a nerve impulse between two cells, and during synaptic transmission the amplitude and frequency of the signal can be adjusted. Some synapses cause depolarization of the neuron, others cause hyperpolarization; the former are excitatory, the latter are inhibitory. Typically, stimulation from several excitatory synapses is necessary to excite a neuron.
The term was introduced in 1897 by the English physiologist Charles Sherrington.
Classification. Structural classification
Based on the number and arrangement of dendrites and axons, neurons are divided into axonless neurons, unipolar neurons, pseudounipolar neurons, bipolar neurons, and multipolar (many dendritic arbors, usually efferent) neurons.
Axonless neurons are small cells grouped near the spinal cord in the intervertebral ganglia, which do not have anatomical signs of division of processes into dendrites and axons. All processes of the cell are very similar. The functional purpose of axonless neurons is poorly understood.
Unipolar neurons - neurons with one process, are present, for example, in the sensory nucleus of the trigeminal nerve in the midbrain. Many morphologists believe that unipolar neurons do not occur in the body of humans and higher vertebrates.
Bipolar neurons are neurons with one axon and one dendrite, located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia.
Multipolar neurons are neurons with one axon and several dendrites. This type of nerve cells predominates in the central nervous system.
Pseudounipolar neurons are unique in their kind. One process extends from the body, which immediately divides in a T-shape. This entire single tract is covered with a myelin sheath and is structurally an axon, although along one of the branches the excitation goes not from, but to the body of the neuron. Structurally, dendrites are branches at the end of this (peripheral) process. The trigger zone is the beginning of this branching (that is, it is located outside the cell body). Such neurons are found in the spinal ganglia.
Neuron(neurocyte), neuronum(neurocytus), has a body, a corpus, a long process-axon, axon, and short branching processes-dendrites, dendrite.
Neurons form chains that transmit a signal - a nerve impulse - from the dendrites to the body and further to the axon, which, branching, contacts the bodies of other neurons, their dendrites or axons. The connection of neurons is carried out through the contact zone - synapse, providing transmission of nerve impulses.
Chemical mediators usually take part in this transmission. When transmitting an impulse, there is a slight delay in the passage of the impulse. Over the course of a person's life, synapses can be destroyed and new synapses can be formed. Memory mechanisms, in particular, are associated with the formation of new contacts between neurons.
Chains of neurons, including an afferent neuron, whose dendrites have sensory endings in various organs, and an efferent neuron, whose axon ends in the working organ (muscle, gland), are designated as the simplest reflex arcs. Typically, in a reflex arc, an impulse is transmitted from a sensitive neuron to an intercalary (associative neuron), and from the latter to an efferent (effector) neuron.
Numerous connections of the associative neuron include the reflex arc in complex neural complexes.
The nervous system develops from the outer germ layer, the ectoderm. The anlage of the nervous system has the form of a neural plate, which is a thickening of the ectoderm along the dorsal surface of the body. Subsequently, the edges of the neural plate, becoming thicker, come closer to each other, while the plate itself, deepening, forms a neural groove. The edges of the plate, taking the form of neural folds, connect and form a neural tube, which, plunging into the depths, is detached from the ectoderm.
At the same time, nodal (ganglionic) plates are formed from the cells that make up the neural folds. Subsequently, they split: part of them, located in the form of ridges on the sides of the neural tube, closer to its dorsal surface, forms spinal nodes; the other part of the nerve cells migrates to the periphery, forming nodes of the autonomic nervous system.
Various differentiation and uneven growth of the neural tube significantly change its internal structure, appearance and shape of the cavity.
The expanded cranial portion of the neural tube develops into brain, and the rest of it into the spinal cord.
Neural tube cells differentiate into neuroblasts, which form neurons with their processes, and into spongioblasts, which give rise to neuroglial elements.
Neurons develop as highly specialized cells. Through their processes, some neurons establish connections between different parts of the brain - this is intercalary (associative) neurons, others communicate the nervous system with other organs - these are afferent (receptor) And efferent (effector) neurons.
The axons of afferent and efferent neurons are part of the nerves that extend from the brain and spinal cord.